【论著】| TCF7转录激活MACC1调节有氧糖酵解促进直肠癌奥沙利铂耐药
时间:2024-09-26 16:00:45 热度:37.1℃ 作者:网络
[摘要] 背景与目的:直肠癌是发生在直肠内壁的癌症,结肠癌转移相关基因-1(metastasis-associated in colon cancer 1,MACC1)能促进结肠癌细胞的耐药性。本研究旨在探究MACC1在直肠癌中对奥沙利铂耐药的影响及机制。方法:分析MACC1和转录因子7(transcription factor 7,TCF7)在直肠癌组织中的表达及MACC1富集的信号转导通路。采用实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)检测直肠癌细胞及直肠癌奥沙利铂耐药细胞中MACC1和TCF7的表达,采用细胞计数试剂盒(cell counting kit-8,CCK-8)检测细胞活力,采用细胞集落形成实验检测细胞的增殖情况,采用Seahorse Biosciences XF96检测ECAR值、乳酸生成量和葡萄糖消耗量,采用蛋白质印迹法(Western blot)检测糖酵解相关蛋白的表达,采用双荧光素酶报告基因和ChIP验证MACC1与TCF7之间的调控关系。采用GraphPad Prism 8.0对各组数据进行正态性检验与方差齐性分析,根据结果是否为正态分布进行单因素方差分析、t检验或Wilcoxon秩和检验。采用双侧检验,P<0.05为差异有统计学意义。结果:MACC1和TCF7在直肠癌中表达上调,敲低MACC1显著抑制了直肠癌耐药细胞活力及不同浓度奥沙利铂处理后的IC50值,也降低了直肠癌奥沙利铂耐药细胞的增殖能力。过表达MACC1能够促进直肠癌细胞增殖和糖酵解能力。进一步研究发现,MACC1上游存在转录因子TCF7,本研究采用癌症基因组图谱计划(The Cancer Genome Atlas Program,TCGA)数据集分析了TCF7在直肠癌组织中的表达量,结果显示,敲低TCF7能够减弱MACC1对直肠癌细胞糖酵解能力及奥沙利铂耐药的促进作用。结论:TCF7/MACC1轴促进有氧糖酵解进而促进直肠癌细胞奥沙利铂耐药,提示靶向TCF7/MACC1轴或抑制有氧糖酵解途径可能是抑制直肠癌奥沙利铂耐药性的新的治疗方法。
[关键词] MACC1;TCF7;糖酵解;直肠癌;奥沙利铂耐药
[Abstract] Background and purpose: Rectal cancer occurs on the inner wall of the rectum, and metastasis-associated in colon cancer 1 (MACC1) can promote drug resistance in colon cancer cells. This study aimed to investigate the effect of MACC1 on oxaliplatin resistance in rectal cancer and its mechanism. Methods: Biosignal analysis of MACC1 and TCF7 expressions in rectal cancer tissues and signaling pathways enriched for MACC1 were carried out. The expressions of MACC1 and TCF7 in rectal cancer cells and rectal cancer oxaliplatin-resistant cells were detected by real-time fluorescence quantitative polymerase chain reaction (RTFQ-PCR), cell viability was detected by cell counting kit-8 (CCK-8), and cell proliferation was detected by cell colony formation assay. ECAR values, lactate production and glucose consumption were detected by Seahorse Biosciences XF96. Western blot was used to detect the expressions of glycolysis-related proteins. Dual luciferase reporter gene and ChIP were used to validate the regulatory relationship between MACC1 and TCF7. Normality test and homogeneity of variance analysis was performed on the data of each group by GraphPad Prism 8.0. If it conformed to normal distribution, one-way ANOVA or t-test would be used for inter group comparison, otherwise the comparison between groups would be conducted using Wilcoxon rank sum test. Using a two-sided test, P<0.05 was considered statistically significant. Results: MACC1 and TCF7 expressions were upregulated in rectal cancer, and knockdown of MACC1 significantly inhibited the viability of rectal cancer drug-resistant cells and the IC50 values of different concentrations of oxaliplatin treatments, as well as reduced the proliferation ability of rectal cancer oxaliplatin-resistant cells. Overexpression of MACC1 was able to promote the proliferation and glycolytic capacity of rectal cancer cells. Further studies revealed that the transcription factor TCF7 existed in the upper reaches of MACC1. Besides, this study analyzed the data from Cancer Genome Atlas Program (TCGA), and the result showed that knockdown of TCF7 was able to attenuate the promotional effect of overexpression of MACC1 on the glycolytic capacity and oxaliplatin resistance of rectal cancer cells. Conclusion: This study demonstrated that the TCF7/MACC1 axis could promote aerobic glycolysis and thus oxaliplatin resistance in rectal cancer cells. The findings suggest that targeting the TCF7/MACC1 axis or inhibiting the aerobic glycolysis pathway may be a novel therapeutic approach to inhibit oxaliplatin resistance in rectal cancer.
[Key words] MACC1; TCF7; Glycolysis; Rectal cancer; Oxaliplatin resistance
最新国家癌症统计数据[1-2]显示,中国结直肠癌病例中结肠癌发病率与直肠癌相当,但在结直肠癌中的比例在不断下降。手术是早期直肠癌的重要治疗手段。近年来,靶向治疗、免疫治疗药物的不断涌现,已广泛用于晚期直肠癌的治疗等[3-4]。目前化疗已被广泛用于改善进展期直肠癌患者的预后[5-6]。然而,化疗过程中,耐药情况的出现导致疾病进展,进而影响患者的预后[7]。因此,解决化疗耐药对改善直肠癌患者的预后有重要意义。
有研究[8]表明,代谢重编程与直肠癌发展有关。糖酵解(又称Warburg效应)能促进癌症转移并重塑肿瘤微环境[9]。Lin等[10]的研究发现,POU2F1高表达能够促进结肠癌细胞的增殖、有氧糖酵解和磷酸戊糖途径,且POU2F1能够通过POU2F1/ALDOA(醛缩酶A)轴来促进代谢重编程和引起奥沙利铂耐药。虽然有氧糖酵解与奥沙利铂耐药有关,但其对直肠癌细胞奥沙利铂耐药的影响及其可能涉及的机制尚不完全清楚。
结肠癌转移相关基因-1( metastasis-associated in colon cancer 1,MACC1)是临床实践中重要的预后标志物,参与多种肿瘤的复发、转移[11]。MACC1导致化疗耐药。Zhang等[12]在肺癌中分析了MACC1的表达水平与顺铂耐药性的关系,结果发现,下调MACC1表达能够抑制肺癌对顺铂耐药。Duan等[13]研究发现,MACC1作为胃癌进展和预后相关的癌基因,可通过调节FANS的表达来驱动胃癌细胞对奥沙利铂耐药。有研究[14-15]报道,MACC1在结直肠癌中呈高表达,且通过β-catenin信号转导通路促进结直肠癌的增殖,但MACC1调控直肠癌细胞进展的相关机制仍需探索。本研究旨在探究MACC1在直肠癌中对奥沙利铂耐药的影响及机制。
1 材料和方法
1.1 材料
1.1.1 实验细胞
直肠癌细胞株Caco-2、SW480、SW837、人正常直肠上皮细胞系FHC以及人胚肾细胞系HEK293T均购自美国典型培养物保藏中心(American Type Culture Collection,ATCC)。
1.1.2 试剂与仪器
二甲基亚砜(dimethyl sulfoxide,DMSO)购自美国Selleckchem公司,奥沙利铂购自美国Sigma公司,oe-MACC1、oe-转录因子7(transcription factor 7,TCF7)及其阴性对照试剂购自深圳华大基因股份有限公司,PolyFast Transfection Reagent试剂和抑制剂2-脱氧-D-葡萄糖(2-DG)购自美国MedChemExpress公司,TRIzol试剂购自美国赛默飞世尔科技公司,Primescript RT预混试剂盒和TB Green® Premix Ex TaqTM购自日本Takara公司,二辛可宁酸(bicinchoninic acid,BCA)检测试剂盒购自美国ThermoFisher Scientific公司,增强型化学发光试剂盒和兔抗人MACC1抗体购自美国Milipore公司,兔抗人β-actin、兔抗人HK2、兔抗人TCF7抗体以及Simple ChIP®Enzymatic Chromatin IP试剂盒购自美国Cell Signaling Technology公司,辣根过氧化物酶标记山羊抗兔IgG(H+L)购自英国Abcam公司,细胞计数试剂盒(cell counting kit-8,CCK-8)购自日本Dojindo公司,Seahorse XF糖酵解压力测试试剂盒购自美国Agilent公司,葡萄糖摄取比色测定试剂盒和乳酸测定试剂盒购自北京索莱宝科技有限公司,PolyFast Transfection Reagent试剂购自美国MedChemExpress公司。
Applied BiosystemsTM7500实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)仪、 Multiskan FC酶标仪购自美国ThermoFisher Scientific公司,Seahorse XF96细胞外通量分析仪购自美国Agilent公司,荧光及化学发光成像仪器购自中国上海勤翔科学仪器有限公司。
1.2 生物信息学分析
本研究中使用癌症基因组图谱计划(The Cancer Genome Atlas Program,TCGA)下载直肠癌中所有的差异表达基因进行差异分析(normal:10, tumor:167),之后结合文献确定研究的目的基因为MACC1和TCF7(|logFC|>1,FDR<0.05)[13]。使用Gene Set Enrichment Analysis分析MACC1所在的基因集所富集通路。使用转录因子motif数据库JASPAR预测与MACC1结合的转录因子,并进行motif位点分析。
1.3 细胞培养
人直肠癌细胞株Caco-2、SW480、SW837、人正常直肠上皮细胞系FHC以及人胚肾细胞系HEK293T均购自美国ATCC,Caco-2细胞在Eagle’s最低必需培养基(minimum essential medium,MEM)中培养,SW480和SW837细胞在Leibovitz’s L-15培养基中培养,FHC细胞在DMEM/F-12培养基中培养,HEK293T细胞在Dulbecco’s modified Eagle’s medium(DMEM)培养基中培养,培养基中均含有10%的胎牛血清(fetal bovine serum,FBS)和1%的青霉素及链霉素混合液,均在37 ℃、CO2体积分数为5%的细胞培养箱中培养。
使用DMSO溶解的奥沙利铂逐渐增加浓度(0、0.5、1.0、1.5和2.0 μmol/L)持续处理直肠癌细胞5个月,进而构建奥沙利铂耐药细胞系SW837/L-OHP[16]。
1.4 细胞转染
使用PolyFast Transfection Reagent试剂将sh-NC/sh-MACC1、oe-NC/oe-MACC1、sh-NC/oe-TCF7质粒转染至直肠癌细胞内。将糖酵解途径抑制剂2-脱氧-D-葡萄糖(2-DG)溶解在DMSO中,以10 μmol/L的浓度处理直肠癌细胞[17]。
1.5 RNA提取与RTFQ-PCR检测
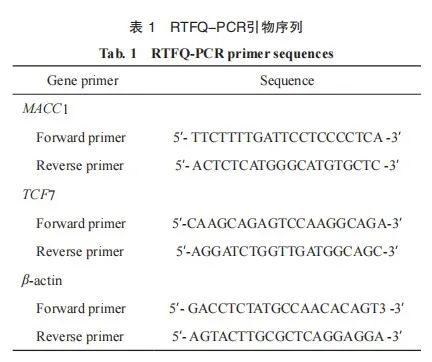
采用TRIzol试剂分离总RNA并进行定量分析。然后根据Primescript RT预混试剂盒反转录合成cDNA,使用TB Green® Premix Ex TaqTM在Applied BiosystemsTM 7500 RTFQ-PCR仪上进行RTFQ-PCR测定。相对目的基因表达以β-actin为内参,使用2-ΔΔCT法进行计算。
1.6 蛋白质印迹法(Western blot)检测
细胞裂解后,使用BCA检测试剂盒测定总蛋白质浓度,将蛋白样本通过十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecylsulphate polyacrylamide gel electrophoresis,SDS-PAGE)进行电泳分离,随后电转移到聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上,用脱脂牛奶(5%)封闭并与一抗温育过夜。然后将膜与辣根过氧化物酶偶联的二抗温育 2 h。使用增强型化学发光试剂盒在荧光及化学发光成像仪器上检测蛋白条带。
1.7 细胞活力与增殖检测
CCK-8试剂盒检测直肠癌细胞奥沙利铂耐药性和耐药细胞的存活率。处理后的SW837/L-OHP细胞铺到培养96孔板中,每孔接5 000个细胞,分别取10 μL CCK-8溶液于培养0、24、48、72 h后添加到每个孔中,37 ℃温育2 h,使用酶标仪检测450 nm波长处的吸光度(D)值。为了评估直肠癌细胞对奥沙利铂的耐药性,在96孔板中每孔接种50 000个细胞,待细胞贴壁后,用递增浓度(0、1.25、2.5、5、10、20和40 μmol/L)的奥沙利铂处理细胞,在48 h后加入10 μL CCK-8溶液,继续温育2 h,使用Multiskan FC酶标仪检测 450 nm波长处的D值,并计算IC50值。
对细胞增殖的检测,将约800个细胞平铺在6孔板中,使用0、1.25、2.5、5、10、20和 40 μmol/L浓度梯度的奥沙利铂处理2周[13]。细胞集落使用4%的多聚甲醛溶液固定并染色。根据细胞集落大小和密集程度评估细胞集落的形成能力。
1.8 细胞外酸化率(extracellular acidification rate,ECAR)值测定
ECAR值使用Seahorse XF糖酵解压力测试试剂盒在Seahorse XF 96细胞外通量分析仪测定。以每孔1×104个细胞接种于Seahorse XF细胞培养板中,基线测量后,依次在各指定时间点加入葡萄糖(glucose)、氧化磷酸化抑制剂寡霉素(oligomycin)、糖酵解抑制剂2-DG来检测ECAR值,数据通过Seahorse XF-96 Wave 软件进行评估,以mpH/min表示。
1.9 葡萄糖摄取、乳酸测定
采用葡萄糖摄取比色测定试剂盒和乳酸测定试剂盒分别测定葡萄糖摄取和乳酸产生量。简而言之,裂解细胞后,使用BCA检测试剂盒测定总蛋白质浓度,根据制造商说明书操作步骤,分别测定葡萄糖摄取和乳酸产生量。
1.10 双荧光素酶活性测定
由上海吉凯基因医学科技股份有限公司构建含有野生型(wild type,WT)3’UTR和突变型(mutant type,MUT)3’UTR的MACC1启动子序列的pmirGLO荧光素酶报告质粒。使用PolyFast Transfection Reagent试剂将上面合成的质粒转染至HEK293T细胞中。48 h后,使用酶标仪器进行荧光素酶活性检测并出具报告。
1.11 ChIP检测
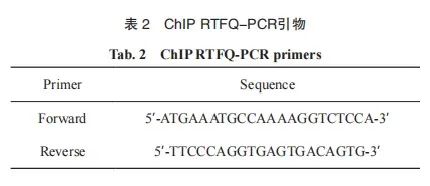
使用Simple ChIP® Enzymatic Chromatin IP Kit检测MACC1与TCF7的结合关系。将细胞用1%多聚甲醛固定10 min,然后加入0.125 mol/L甘氨酸,并将混合物在室温下放置5 min以终止DNA-蛋白质交联。向细胞中加入SDS裂解液(需加蛋白酶抑制剂),使用超声破碎仪使染色质片段化。将裂解液与兔抗TCF7抗体和Protein G磁珠温育,形成DNA-抗体-磁珠复合物,洗脱纯化后标记为Target。兔抗IgG 抗体作为阴性对照。通过RTFQ-PCR对最终纯化的DNA片段进行分析。MACC1启动子结合位点的RTFQ-PCR引物见表2。
1.12 统计学处理
将各项数据以3次独立重复的均数以x±s表示。对各组数据进行正态性检验与方差齐性分析,如符合正态分布且方差齐,则组间比较采用单因素方差分析或t检验;如不符合,则组间比较采用Wilcoxon秩和检验。采用双侧检验,P<0.05为差异有统计学意义。所有的统计分析均采用GraphPad Prism 8.0(GraphPad software,USA)软件进行。
2 结 果
2.1 MACC1在直肠癌中表达上调
本研究在TCGA数据库中分析了直肠癌中的MACC1的表达。结果发现,在直肠癌组织中,MACC1的表达显著上调(图1A)。对直肠癌细胞系也进行了MACC1的表达分析,发现与组织中分析的结果一致,MACC1在直肠癌细胞中的表达均上调(图1B),但分布情况均不符合正态分布。经分析试验数据表明,MACC1在直肠癌中呈高表达。
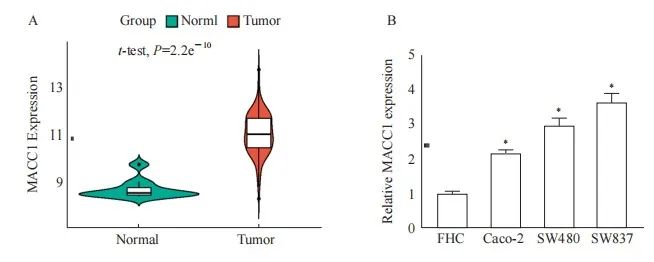
图1 MACC1在直肠癌中表达上调
Fig. 1 Upregulation of MACC1 expression in rectal cancer
A: Analyze the expression level of MACC1 in normal tissues adjacent to cancer tissues and rectal cancer tissues using the TCGA database; B: RTFQ-PCR analysis of MACC1 expression in Caco-2, SW480, SW837, and normal rectal epithelial cells FHC. *: P<0.05, compared with each other.
2.2 MACC1促进直肠癌细胞奥沙利铂耐药
挑选表达量较高的细胞系SW837构建奥沙利铂耐药直肠癌细胞系(SW837/L-OHP)。RTFQ-PCR结果显示,在奥沙利铂耐药细胞系中MACC1的表达显著高于直肠癌奥沙利铂敏感细胞系(图2A)。随后将sh-MACC1转染至SW837/L-OHP细胞系中,结果发现,MACC1在SW837/L-OHP细胞系中MACC1的表达显著低于对照组(图2B)。此外,敲低MACC1的耐药细胞的活力也显著低于对照组细胞的活力(图2C)。与对照相比,sh-MACC1组直肠癌奥沙利铂耐药细胞(SW837/L-OHP)的增殖能力明显减弱(图2D、E)。随后使用不同浓度的奥沙利铂处理SW837/L-OHP,CCK-8检测结果发现,与对照组相比,敲低MACC1后SW837/L-OHP细胞对奥沙利铂敏感(图2F)。
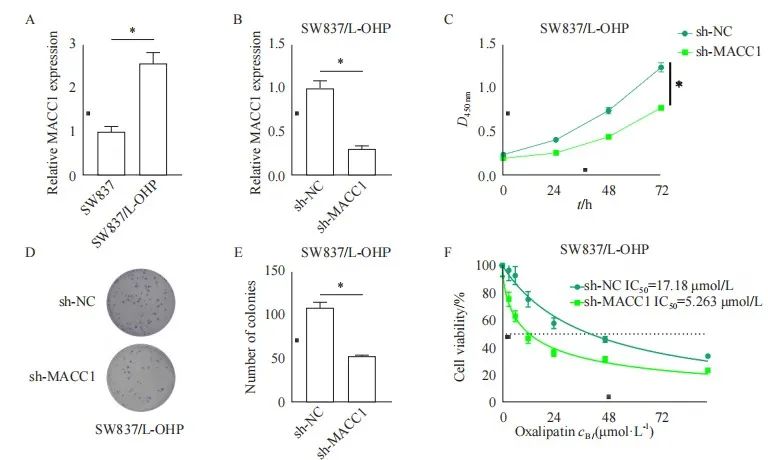
图2 MACC1促进直肠癌细胞的耐药
Fig. 2 MACC1 promotes drug resistance in rectal cancer cells
A: The expression of MACC1 in oxaliplatin sensitive rectal cancer cell lines and oxaliplatin resistant rectal cancer cell lines; B: Detecting the transfection efficiency of sh-MACC1; C: Detection of the activity of rectal cancer oxaliplatin resistant cell lines after knocking down MACC1; D-E: Detection of decreased proliferation ability of rectal cancer oxaliplatin resistant cells after knocking down MACC1; F: Detection of the IC50 value of rectal cancer oxaliplatin resistant cells after knocking down MACC1. *: P<0.05, compared with each other.
2.3 MACC1通过促进有氧糖酵解促进直肠癌细胞奥沙利铂耐药
本研究通过生物信息学方法分析了MACC1具体参与调控直肠癌细胞奥沙利铂耐药的机制。结果显示,MACC1富集在GLYCOLYSIS GLUCONEOGENESIS通路上,且与糖酵解标志基因的转录和表达呈正相关(图3A、B)。有研究[18]报道,糖酵解是诱导肿瘤耐药的重要因素,因此我们探索了直肠癌细胞中的糖酵解能力。与oe-NC+DMSO相比,oe-MACC1+DMSO组的SW837/L-OHP细胞的活力显著增加,而加入2-DG后则使过表达MACC1后的细胞活力恢复到正常水平(图3C)。为了进一步探索MACC1如何调节糖酵解重编程,我们测定了直肠癌SW837/L-OHP细胞中的ECAR值、葡萄糖消耗量和乳酸生成量。结果表明,过表达MACC1可显著提高SW837/L-OHP细胞的ECAR值、乳酸生成量以及葡萄糖消耗量,而进一步添加2-DG则使SW837/L-OHP细胞的ECAR值、乳酸生成量以及葡萄糖消耗量降低(图3D、E)。同样,使用Western blot检测糖酵解通路中的关键蛋白表达,结果发现,过表达MACC1后HK2表达明显上调,加入2-DG的同时过表达MACC1处理的SW837/L-OHP细胞的HK2蛋白表达又恢复到对照组水平(图3F)。此外,2-DG加入后部分减弱了过表达MACC1后SW837/L-OHP细胞增殖能力的促进作用(图3G)。不同浓度的奥沙利铂处理SW837/L-OHP细胞后,过表达MACC1组的IC50值明显升高,而加入2-DG后,过表达MACC1处理组的SW837/L-OPH细胞的IC50值恢复到正常水平(图3H)。上述结果表明,MACC1可通过促进有氧糖酵解进而促进直肠癌细胞对奥沙利铂耐药。
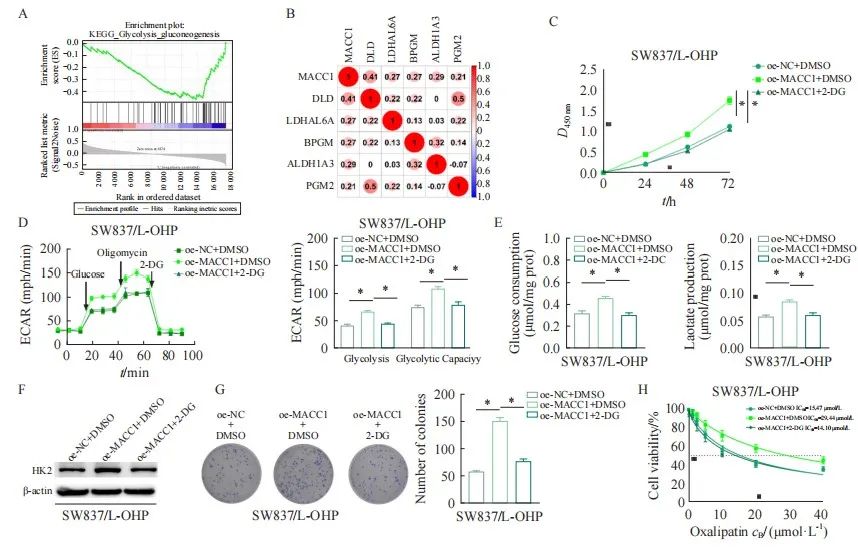
图3 MACC1通过调节有氧糖酵解促进直肠癌细胞奥沙利铂耐药
Fig. 3 MACC1 promotes oxaliplatin resistance in rectal cancer cells by regulating aerobic glycolysis
A: GESA analysis revealed that MACC1 was enriched in glycolytic signaling pathways; B: The correlation between MACC1 and glycolytic marker genes; C: Detect the cell viability of rectal cancer oxaliplatin resistant cells after expressing MACC1 and adding 2-DG; D: Detect the ECAR value of rectal cancer cells after oe-MACC1+DMSO; E: Glucose consumption and lactate production in rectal cancer oxaliplatin resistant cells after oe-MACC1+DMSO treatment; F: Detecting the expression of key protein HK2 in the glycolytic pathway; G: Colony formation assay was used to detect the proliferation ability of rectal cancer oxaliplatin resistant cells treated with oe-MACC1 and oe-MACC1+2-DG; H: Detect the IC50 values of rectal cancer oxaliplatin resistant cells treated with different concentrations of oxaliplatin.*: P<0.05, compared with each other.
2.4 TCF7转录激活MACC1
为了进一步探究MACC1上游调控机制,本研究通过ChIPBase预测了MACC1启动子区域的转录因子。结果发现,一共存在68个转录因子(图4A)。通过筛选分析发现TCF7是其中的一个转录因子,与MACC1表达存在正相关的关系(图4B)。因此我们使用TCGA-数据集分析了TCF7在直肠癌组织中的表达量,发现TCF7在直肠癌组织中高表达(图4C)。随后在直肠癌细胞以及直肠癌奥沙利铂耐药细胞SW837/L-OHP中验证了TCF7的表达水平,且与TCGA预测结果一致,表达均上调(图4D)。随后,我们通过JASPAR分析确定了TCF7与MACC1启动子区域的结合位点(图4E),ChIP和双荧光素报告基因实验结果显示,相较于IgG组,MACC1在抗TCF7分组细胞中被显著富集;此外,与对照组中相比,sh-TCF7能够显著降低MACC1-WT的荧光素酶活性,对MACC1-MUT没有影响;以上结果显示,TCF7能够转录调控下游MACC1的表达(图4F)。
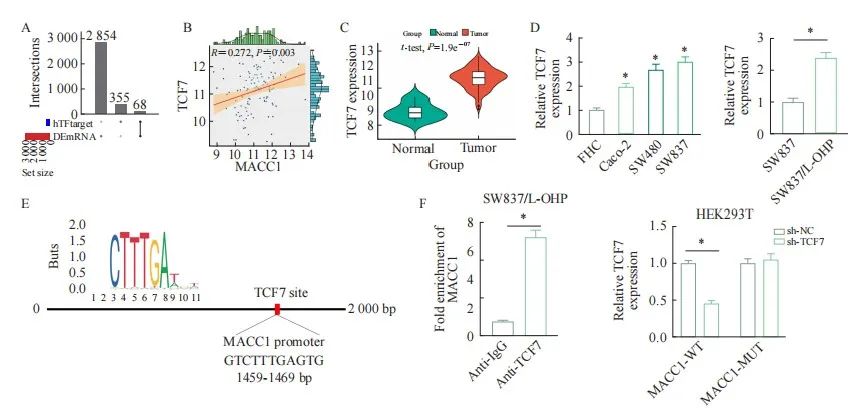
图4 TCF7转录调控MACC1
Fig. 4 TCF7 transcription regulation of MACC1
A: ChIPBase predicted transcription factors that bind to the promoter region of MACC1; B: There was a positive correlation between TCF7 and MACC1; C: The expression level of TCF7 in rectal cancer tissue; D: The expression level of TCF7 in rectal cancer cells and oxaliplatin resistant cells; E: The binding motif of TCF7 and MACC1; F: ChIP and dual luciferase assays were performed to detect the key binding points of MACC1 and TCF7 in SW837/L-OHP cells.*: P<0.05, compared with each other.
2.5 TCF7靶向MACC1调节有氧糖酵解促进直肠癌细胞奥沙利铂耐药
由于MACC1转录受TCF7调控,本研究继而深入探讨了直肠癌奥沙利铂耐药细胞SW837/L-OHP中TCF7转录激活MACC1调节有氧糖酵解对直肠癌细胞奥沙利铂耐药的影响。通过检测构建的耐药细胞系中敲低TCF7、敲低TCF7并过表达MACC1、过表达TCF7以及敲低TCF7和MACC1后各组的转染效率和细胞活力,结果表明,敲低TCF7后MACC1的表达下调,敲低TCF7后直肠癌奥沙利铂耐药细胞活力也下降,而敲低TCF7的同时进一步过表达MACC1后,MACC1的表达恢复到对照组水平,且细胞活力的降低也得到缓解。过表达TCF7后MACC1的表达上调,而过表达TCF7的同时敲低MACC1后,MACC1的表达情况恢复到对照组水平,且细胞活力的上升也得到缓解(图5A、B)。检测各组细胞的糖酵解能力发现,敲低TCF7后,ECAR降低,乳酸生成量和葡萄糖消耗量下降;过表达TCF7后, ECAR降低,乳酸生成量和葡萄糖消耗量上升;敲低TCF7并过表达MACC1、过表达TCF7并敲低MACC1,ECAR值、葡萄糖消耗量及乳酸生成量恢复到相应的对照组水平(图5C~E)。不仅如此,在敲低TCF7后,糖酵解相关蛋白HK2的表达下调,过表达TCF7后,糖酵解相关蛋白HK2的表达上调;而在敲低TCF7的直肠癌奥沙利铂耐药细胞系中同时过表达MACC1、过表达TCF7的直肠癌奥沙利铂耐药细胞系中同时敲低MACC1后,HK2的表达恢复到相应的对照组的水平(图5F)。此外,我们发现过表达MACC1显著阻断了敲低TCF7对SW837/L-OHP细胞的集落形成能力的抑制作用。敲低MACC1显著增强了过表达TCF7对SW837/L-OHP细胞的集落形成能力(图5G)。并且不同浓度的奥沙利铂处理SW837/L-OHP细胞后,敲低TCF7使得直肠癌奥沙利铂耐药细胞对奥沙利铂更加敏感,过表达TCF7使得直肠癌奥沙利铂耐药细胞对奥沙利铂不敏感。但是在转染sh-TCF7+oe-MACC1组我们观察到直肠癌奥沙利铂耐药细胞的敏感性下降,对应的oe-TCF7+sh-MACC1组直肠癌奥沙利铂耐药细胞的敏感性上升(图5H)。上述结果表明,TCF7调控MACC1促进有氧糖酵解途径影响直肠癌细胞奥沙利铂耐药性。
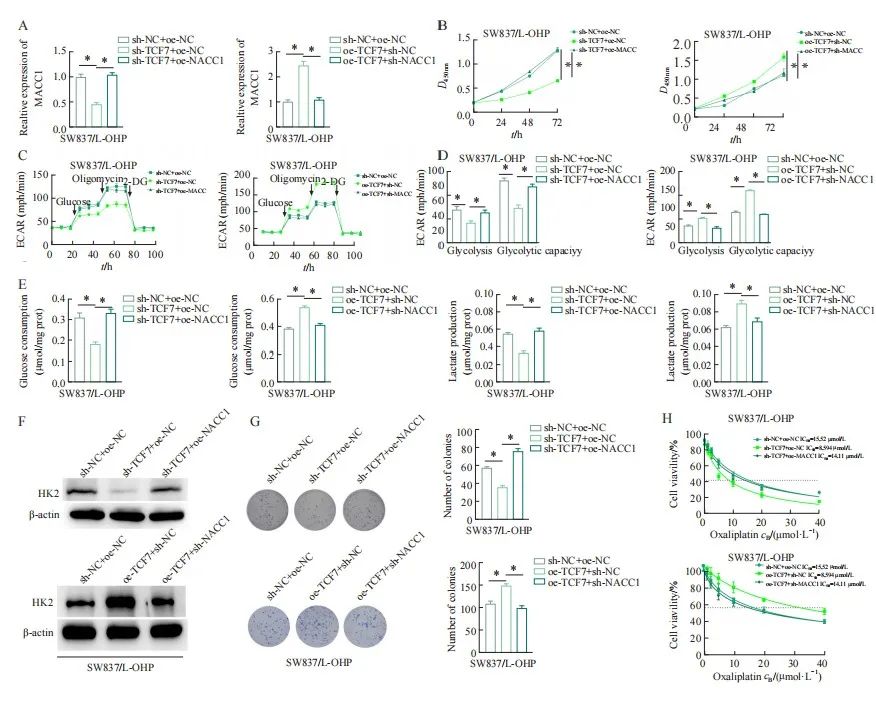
图5 TCF7/MACC1调节有氧糖酵解途径影响直肠癌细胞奥沙利铂耐药
Fig. 5 TCF7/MACC1 regulated aerobic glycolysis pathway and affects oxaliplatin resistance in rectal cancer cells
A: Detection of transfection efficiency in rectal cancer oxaliplatin-resistant cells transfected with knockdown TCF7 plasmid, knockdown TCF7 and overexpressing MACC1 plasmid, transfected with overexpression of TCF7 plasmid, overexpression of TCF7 and knockdown of MACC1 plasmid rectal cancer oxaliplatin-resistant cells; B: Transfection of rectal cancer oxaliplatin-resistant cells with knockdown TCF7 plasmid, knockdown TCF7 and overexpression of MACC1 plasmid, cell viability of rectal cancer oxaliplatin-resistant cells transfected with overexpression of TCF7 plasmid, overexpression of TCF7 and knockdown of MACC1 plasmid; C-D: Detection of ECAR values of rectal cancer oxaliplatin-resistant cells in each treatment group; E: Glucose consumption as well as lactic acid production of rectal cancer oxaliplatin-resistant cells in each treatment group; F: Western blot assay for the expression of key proteins in the glycolytic pathway; G: SW837/L -OHP cells colony forming ability under different treatments; H: IC50 values of SW837/L-OHP cells under different concentrations of oxaliplatin; *: P<0.05, compared with each other.
3 讨 论
奥沙利铂是已被证明对结直肠癌切实有效的铂类药物,并已成为该肿瘤的一线化疗方案用药,不仅如此,奥沙利铂也应用于胰腺癌及胃癌的辅助治疗[19-20]。然而,此类肿瘤患者常出现治疗失败,主要是由于获得性耐药[21]。因此,了解奥沙利铂的耐药机制对于提高结直肠癌的治疗效果非常重要。对于奥沙利铂耐药机制的最近研究进展,包括药物摄入、转出、DNA损伤修复、细胞死亡、自噬在调控奥沙利铂耐药中的关键作用[29-31]。因此,研究者们针对顺铂作用及其耐药机制和新的铂类药物进行了大量研究。前人研究表明MACC1是治疗结直肠癌奥沙利铂耐药的重要靶点,一项关于靶向结直肠癌细胞中的MACC1的研究[25]表明,FOXA3/MACC1轴能够破坏结直肠癌肿瘤细胞的干性并调节结直肠癌细胞对奥沙利铂的敏感性。另有研究[26]发现,结直肠癌细胞内的循环RNA超氧化物歧化酶(circular RNA copper chaperone for superoxide dismutase,circ-CCS)可通过靶向miR-874-3P/HK2轴调控糖酵解途径进而影响奥沙利铂的耐药性。MACC1已被发现存在于20多种不同类型的实体瘤中,可作为治疗和预防癌症的潜在靶点[27]。
Zhang等[12]的研究结果表明,在顺铂耐药的肺癌细胞系中,MACC1的表达水平升高,并且肺癌细胞随着MACC1表达的降低对顺铂的敏感性增强。虽然Zhang等[12]的研究结果已表明MACC1与肿瘤耐药相关,但其是否参与直肠癌奥沙利铂耐药的具体机制还不清楚。本研究的结果表明,MACC1在直肠癌组织、奥沙利铂耐药细胞中均呈高表达。此外,本研究发现,敲低MACC1能够抑制SW837/L-OHP细胞的IC50值,这意味着MACC1可能参与直肠癌中奥沙利铂耐药过程。Li等[28]发现,MACC1高表达预示着胃癌切除术后的高复发率,低表达MACC1能够抑制糖酵解代谢进而抑制胃癌细胞的增殖,而糖酵解能力的上调能够促进结直肠癌细胞对奥沙利铂耐药的发生[29]。本研究的结果也发现,MACC1富集在GLYCOLYSIS GLUCONEOGENESIS信号转导通路上,并且MACC1与有氧糖酵解标志基因的表达呈正相关。总之,本研究的结果表明, MACC1可通过促进糖酵解途径抑制直肠癌肿瘤对奥沙利铂的敏感性,这一发现为靶向糖酵解代谢途径治疗奥沙利铂耐药的直肠癌提供了依据。
本研究进一步分析MACC1在直肠癌奥沙利铂耐药性中发挥促进作用具体调控机制,结果发现,TCF7与MACC1的启动子区域具有结合关系,且TCF7能够转录激活MACC1的表达。TCF7是TCF家族的成员,具有TCF家族的分子特征,即高迁移率基团DNA结合结构域和β-catenin结合结构域[30]。作为一种转录激活因子,多项研究[31-33]显示,TCF7过表达会促进多种肿瘤的进展。本研究对TCF7在直肠癌中的表达进行了验证,结果发现,TCF7呈高表达,并且TCF7是MACC1的上游调控分子。此外,我们发现TCF7能够转录激活MACC1促进糖酵解途径进而促进直肠癌细胞对奥沙利铂的耐药性。上述结果表明,靶向TCF7/MACC1轴或抑制糖酵解途径可能是治疗奥沙利铂耐药直肠癌的新手段。
本研究对直肠癌组织及细胞进行检测发现MACC1表达上调,且MACC1高表达能够提升奥沙利铂耐药直肠癌细胞的活力、ECAR值、乳酸产生量、糖酵解相关蛋白HK2的表达水平以及细胞集落形成能力和IC50值,且发现MACC1能够通过促进糖酵解进而促进直肠癌细胞对奥沙利铂耐药。同时,TCF7能够转录激活MACC1,且TCF7/MACC1轴能够促进糖酵解进而促进直肠癌细胞对奥沙利铂的耐药性。本研究结果表明, MACC1在直肠癌组织及耐药细胞中明显高表达,TCF7与MACC1的表达呈正相关。体外研究结果显示,TCF7/MACC1轴通过糖酵解代谢途径进而促进直肠癌细胞对奥沙利铂耐药。
本研究尚存在如下不足:首先,仅基于体外实验得到以上结果,尚需要通过动物实验进行验证;其次关于MACC1调节糖酵解途径的关键酶有待揭示。总之,本研究结果显示,靶向TCF7/MACC1来调节糖酵解途径可能是逆转直肠癌奥沙利铂耐药的新的治疗策略。
无利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:
严研:确定选题,设计并实施研究,分析数据,撰写文章;周立庆:提供研究思路,分析研究方案可行性,参与文章的修订与审核;夏建洪:参与提出文章的撰写思路,收集并分析数据,参与文章的修订;马婷婷:收集并分析数据,设计论文框架,参与文章的修订及终版文章的审核。
[参考文献]
[1] ZHENG R S, ZHANG S W, ZENG H M, et al. Cancer incidence and mortality in China, 2016[J]. J Natl Cancer Cent, 2022, 2(1): 1-9.
上海市抗癌协会大肠癌专业委员会. 结直肠癌早筛、早诊、早治上海方案(2023年版)[J]. 中国癌症杂志, 2024, 34(1): 13-66.
[2] Colorectal Cancer Special Committee of Shanghai Anti-Cancer Association. Shanghai plan for early screening, diagnosis and treatment of colorectal cancer (2023 edition)[J]. China Oncol, 2024, 34(1): 13-66.
[3] YANG Y, WANG H Y, CHEN Y K, et al. Current status of surgical treatment of rectal cancer in China[J]. Chin Med J, 2020, 133(22): 2703-2711.
[4] WANG Y Q, SHEN L J, WAN J F, et al. Neoadjuvant chemoradiotherapy combined with immunotherapy for locally advanced rectal cancer: a new era for anal preservation[J]. Front Immunol, 2022, 13: 1067036.
[5] CIARDIELLO F, CIARDIELLO D, MARTINI G, et al. Clinical management of metastatic colorectal cancer in the era of precision medicine[J]. CA Cancer J Clin, 2022, 72(4): 372-401.
[6] TATARYN B, KRYZHANIVSKA A, HOLOTIUK V, et al. Chemotherapy in rectal cancer[J]. Wiad Lek, 2021, 74(8): 1816-1823.
[7] KIM J K, MARCO M R, ROXBURGH C S D, et al. Survival after induction chemotherapy and chemoradiation versus chemoradiation and adjuvant chemotherapy for locally advanced rectal cancer[J]. Oncologist, 2022, 27(5): 380-388.
[8] ZHANG Z Y, YAO Q Z, LIU H Y, et al. Metabolic reprogramming-associated genes predict overall survival for rectal cancer[J]. J Cell Mol Med, 2020, 24(10): 5842-5849.
[9] LIU C, JIN Y, FAN Z M. The mechanism of Warburg effect induced chemoresistance in cancer[J]. Front Oncol, 2021, 11: 698023.
[10] LIN J G, XIA L Z, OYANG L, et al. The POU2F1-ALDOA axis promotes the proliferation and chemoresistance of colon cancer cells by enhancing glycolysis and the pentose phosphate pathway activity[J]. Oncogene, 2022, 41(7): 1024-1039.
[11] HOHMANN T, HOHMANN U, DEHGHANI F. MACC1- induced migration in tumors: current state and perspective[J]. Front Oncol, 2023, 13: 1165676.
[12] ZHANG Q, ZHANG B, SUN L N, et al. Cisplatin resistance in lung cancer is mediated by MACC1 expression through PI3K/AKT signaling pathway activation[J]. Acta Biochim Biophys Sin, 2018, 50(8): 748-756.
[13] DUAN J M, CHEN L S, ZHOU M Y, et al. MACC1 decreases the chemosensitivity of gastric cancer cells to oxaliplatin by regulating FASN expression[J]. Oncol Rep, 2017, 37(5): 2583-2592.
[14] ZHEN T T, DAI S J, L I H, et al. MACC1 promotes carcinogenesis of colorectal cancer via β-catenin signaling pathway[J]. Oncotarget, 2014, 5(11): 3756-3769.
[15] KORTÜM B, RADHAKRISHNAN H, ZINCKE F, et al. Combinatorial treatment with statins and niclosamide prevents CRC dissemination by unhinging the MACC1-β-catenin S100A4 axis of metastasis[J]. Oncogene, 2022, 41(39): 4446-4458.
[16] CHENG C Y, XIE Z H, LI Y H, et al. PTBP1 knockdown overcomes the resistance to vincristine and oxaliplatin in drug resistant colon cancer cells through regulation of glycolysis[J]. Biomed Pharmacother, 2018, 108: 194-200.
[17] LIU J, PAN C Q, GUO L H, et al. A new mechanism of trastuzumab resistance in gastric cancer: MACC1 promotes the Warburg effect via activation of the PI3K/AKT signaling pathway[J]. J Hematol Oncol, 2016, 9(1): 76.
[18] LI M X, CHE N, JIN Y, et al. CDKN3 overcomes bladder cancer cisplatin resistance via LDHA-dependent glycolysis reprogramming[J]. Onco Targets Ther, 2022, 15: 299-311.
[19] WAINBERG Z A, BEKAII-SAAB T, BOLAND P M, et al. First-line liposomal irinotecan with oxaliplatin, 5-fluorouraciland leucovorin (NALIRIFOX) in pancreatic ductal adenocarcinoma: a phase Ⅰ/Ⅱ study[J]. Eur J Cancer, 2021, 151: 14-24.
[20] PARK S H, LIM D H, SOHN T S, et al. A randomized phase Ⅲ trial comparing adjuvant single-agent S1, S-1 with oxaliplatin, and postoperative chemoradiation with S-1 and oxaliplatin in patients with node-positive gastric cancer after D2 rep: the ARTIST 2 trial [J]. Ann Oncol, 2021, 32(3): 368-374.
[21] KOBA R, FUJITA H, NISHIBORI M, et al. Quantitative evaluation of the intratumoral distribution of platinum in oxaliplatin-treated rectal cancer: in situ visualization of platinum via synchrotron radiation X-ray fluorescence spectrometry[J]. Int J Cancer, 2020, 146(9): 2498-2509.
[22] LI H F, WANG C M, LAN L X, et al. PARP1 inhibitor combined with oxaliplatin efficiently suppresses oxaliplatin resistance in gastric cancer-derived organoids via homologous recombination and the base excision repair pathway[J]. Front Cell Dev Biol, 2021, 9: 719192.
[23] PAN Z H, ZHENG J, ZHANG J B, et al. A novel protein encoded by exosomal CircATG4B induces oxaliplatin resistance in colorectal cancer by promoting autophagy[J]. Adv Sci, 2022, 9(35): e2204513.
[24] LI H, CHEN J H, LIU J, et al. CPT2 downregulation triggers stemness and oxaliplatin resistance in colorectal cancer via activating the ROS/Wnt/β-catenin-induced glycolytic metabolism[J]. Exp Cell Res, 2021, 409(1): 112892.
[25] LI N, LI Y, GAO H B, et al. Forkhead-box A3 (FOXA3) represses cancer stemness and partially potentiates chemosensitivity by targeting metastasis-associated in colon cancer 1 (MACC1) signaling pathway in colorectal cancer cells[J]. Curr Cancer Drug Targets, 2020.
[26] QIU X F, XU Q H, LIAO B L, et al. Circ-CCS regulates oxaliplatin resistance via targeting miR-874-3p/HK2 axis in colorectal cancer[J]. Histol Histopathol, 2023, 38(10): 1145-1156.
[27] LV M M, JIAO Y J, YANG B W, et al. MACC1 as a potential target for the treatment and prevention of breast cancer[J]. Biology, 2023, 12(3): 455.
[28] LIN L, LIU Y T, PAN C Q, et al. Gastric cancer cells escape metabolic stress via the DLC3/MACC1 axis[J]. Theranostics, 2019, 9(7): 2100-2114.
[29] WANG X Y, ZHANG H Y, YANG H O, et al. Exosome delivered circRNA promotes glycolysis t o induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer[J]. Mol Oncol, 2020, 14(3): 539-555.
[30] LIU H, LI D X, SUN L N, et al. Interaction of lncRNA MIR100HG with hnRNPA2B1 facilitates m6A-dependent stabilization of TCF7L2 mRNA and colorectal cancer progression[J]. Mol Cancer, 2022, 21(1): 74.
[31] GUO Y X, GUO Y Y, CHEN C, et al. Circ3823 contributes to growth, metastasis and angiogenesis of colorectal cancer: involvement of miR-30c-5p/TCF7 axis[J]. Mol Cancer, 2021, 20(1): 93.
[32] PENG Y, XIAO L P, RONG H X, et al. Single-cell profiling of tumor-infiltrating TCF1/TCF7+ T cells reveals a T lymphocyte subset associated with tertiary lymphoid structures/organs and a superior prognosis in oral cancer[J]. Oral Oncol, 2021, 119: 105348.
[33] LIU H Q, ZHANG H, LIU X Q, et al. Pancreatic stellate cells exploit Wnt/β-catenin/TCF7-mediated glutamine metabolism to promote pancreatic cancer cells growth[J]. Cancer Lett, 2023, 555: 216040.

