Cell Reports: 少突胶质细胞的巨自噬性髓鞘周转在神经退行性疾病中的作用
时间:2022-11-29 21:02:28 热度:37.1℃ 作者:网络
尽管巨自噬缺陷与神经退行性疾病有关,但我们对中枢神经系统中不连续的、高度进化的细胞类型如何利用巨自噬来维持体内稳态知之甚少。其中有一种细胞类型是少突胶质细胞,其形成的髓鞘是动作电位传导的核心。近期,在《Cell Reports》期刊发表了题为“Oligodendroglial macroautophagy is essential for myelin sheath turnover to prevent neurodegeneration and death”的论文,作者使用小鼠遗传学、活细胞成像、电子显微镜和生物化学等综合方法,表明成熟的少突胶质细胞需要巨自噬的作用,通过在降解前将胞质和跨膜的髓鞘蛋白整合到自噬内涵体,来自主降解细胞的髓鞘。
作者发现,自噬性髓鞘周转的中断会导致髓鞘结构的变化,最终损害神经功能并导致成人发病的进行性运动障碍、神经退行性变和死亡。作者的模型表明,通过巨自噬持续和自主地维持髓鞘功能是非常重要的,这使我们深入了解巨自噬失调如何导致神经退行性疾病的病理生理学。
 成人中枢神经系统髓鞘的持续重塑需要巨自噬的参与
成人中枢神经系统髓鞘的持续重塑需要巨自噬的参与
为了研究巨自噬在髓鞘维持中的作用,作者使用CNP1位点表达的 Cre 重组酶,通过 Atg7 (Atg7cKO) 的纯合性缺失,使少突胶质细胞中的巨自噬失活。正如预期的那样,自噬衔接蛋白 p62 在 Atg7cKO 中积累,但在杂合子 (Atg7cHet) 或 Atg7loxP/loxP (Atg7fl/fl)白质中没有积累。尽管巨自噬完全失活,但 Atg7cKO 小鼠在2 个月大时的生存能力或行为没有明显差异。
髓鞘蛋白的转录不受影响,对前脑白质分析表明不同基因型的少突胶质细胞前体细胞(OPCs)和成熟少突胶质细胞的数量也相似。这些数据表明,在Atg7cKO 小鼠少突胶质细胞系中,CNPCre介导的巨自噬基因失活并不影响少突胶质细胞或髓鞘的形成。在有条件地敲除 Atg5 后获得了类似的结果,Atg5也需要自噬体的形成。
接下来,作者对成人中枢神经系统白质进行了超微结构研究(图1)。尽管2个月大时不同基因型的髓鞘厚度相似,但Atg7cKO小鼠的髓鞘厚度随着年龄的增长而增加(图 1A-1H)。然而,髓磷脂的周转未受影响。对视神经中每个轴突直径的g-ratio分析表明,Atg7cKO 小鼠的g-ratio明显较小,只有最粗的轴突没有明显差异(图 1A-1F)。胼胝体在鞘厚度上显示出类似的显著差异(图1G和1H),并且两个大脑区域也表现出异常髓鞘结构的患病率增加(图1I)。虽然之前的研究表明,在较小的年龄时没有差异,但 Atg7cKO 小鼠在16个月大时的脊髓髓鞘化受到影响,表明自噬在不同的少突胶质细胞中具有类似的重要性。尽管Atg7在外周白质中丢失,但髓鞘形成未受影响,这与之前的报告一致,表明施万细胞重塑髓鞘不需要巨自噬。
然后,作者研究了成年期g-ratio如何变化。分析显示,虽然对照组髓鞘厚度在2到16个月内保持相对不变,但随着巨自噬的破坏,厚度发生了急剧变化(图1J)。这表明少突胶质细胞通过合成和巨自噬介导的降解的平衡来持续维持其髓鞘厚度。此外,这也支持了这样的猜想:即少突胶质细胞中巨自噬的破坏阻碍了体内中枢神经系统髓鞘的重塑。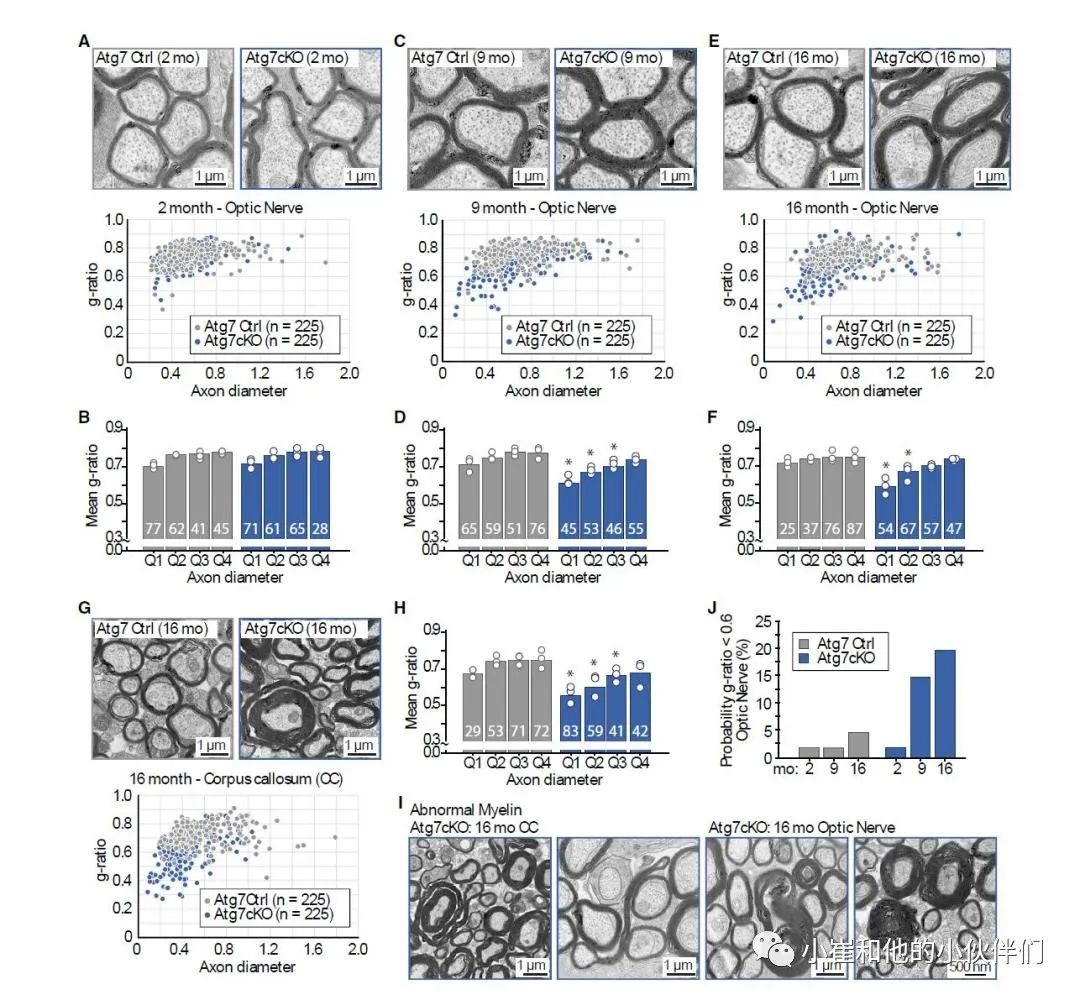 图 1. 少突胶质细胞中自噬的失活导致成年后髓鞘积累、髓鞘厚度增加及髓鞘结构异常
图 1. 少突胶质细胞中自噬的失活导致成年后髓鞘积累、髓鞘厚度增加及髓鞘结构异常
自噬体的形成和成熟可以发生在成熟的少突胶质细胞中
为了证实随着时间的推移,髓鞘厚度的增加是由持续的髓鞘重塑驱动而不是随着时间的推移对自噬活动需求的增加,作者检查了小鼠胼胝体 作为自噬驱动的代表的Olig2+ 细胞中自噬体的存在(图2A 和2B)。用氯喹急性处理GFP(GFP-LC3)标记的表达自噬体标记物 MAP 1A/1B-LC3 的转基因成年小鼠,以减缓自噬体的周转,作者发现,随着小鼠年龄的增长,点状物的强度并没有发生很大变化,如果有的话,还会下降,这表明在不同时期对少突胶质细胞自噬的相对依赖是相当恒定的,并且髓鞘的自噬降解可能在这些细胞中持续发生。
在组织中,自噬体最可能出现在少突胶质细胞体中,因此,为了更好地解释巨自噬如何重塑单个髓鞘,作者通过免疫抗体包被筛选法分离了少突胶质前体细胞,并诱导其分化和成熟。针对Atg16(自噬体生物发生的指标)、p62 和 LC3 的免疫荧光显示,这些蛋白质在少突胶质细胞发育的所有阶段都有表达(图 2C-2H)。重要的是,在成熟的少突胶质细胞中,这些自噬标记物在细胞体和整个髓鞘的细胞质通道中都表达(图2D、2F 和2H)。
为了确认自噬体可以在这些细胞质通道内形成,作者验证了这些细胞质通道内是否存在已知的自噬体膜来源,完整内质网 (ER) 膜蛋白钙联接蛋白的免疫荧光证实了之前的发现,表明 ER 在整个成熟少突胶质细胞中的存在并且自噬体生物发生的潜在膜源存在于髓鞘的细胞质通道中。
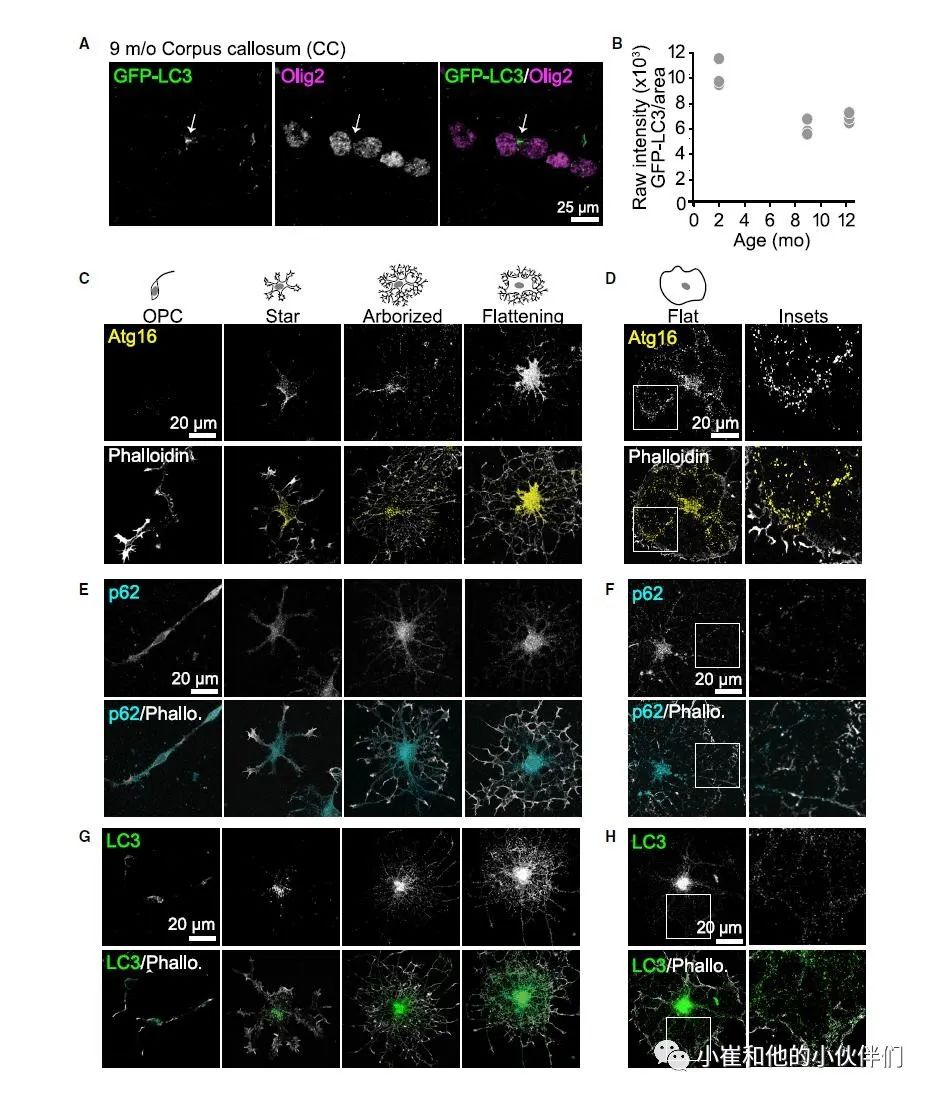 图2. 体内和培养中少突胶质细胞的基础自噬作用
图2. 体内和培养中少突胶质细胞的基础自噬作用
鉴于巨自噬是一个多步骤的程序性事件,作者接下来对培养中成熟的GFP-LC3+少突胶质细胞进行了活细胞转盘式共聚焦显微镜观察。监测和分析细胞新形成的自噬体的出现、酸化(通过LysoTracker红色阳性)以及其消失。作者观察到巨自噬活动的区域异质性,因此设计了一个命名法来描述细胞质通道网络内的亚结构(图 3A):Artery是大通道,可能对应于髓鞘的外环和节旁环;Branch是小通道,可能对应于髓鞘内细胞质纳米通道;Edge是可能对应于内环的通道;Bulb是通道内的扩大部分,通常在动脉的远端发现。
尽管自噬体在酸化时显示出顺向和逆向运动,但与神经元相比,绝大多数结构在整个形成和酸化过程中保持静止(图3B-3D)。自噬体在整个细胞中形成并酸化,大部分存在于髓鞘内的通道内,在Artery-Branch交叉点和Bulb中出现的频率特别高(图 3E)。此外,结构酸化的时间因亚细胞区域而有所不同,并且与细胞体的距离相关,距离较近的酸化速度略快(图3F)。总之,作者的研究表明,自噬体的生物发生和成熟可以发生在成熟的少突胶质细胞中,包括在髓鞘的细胞质通道内。
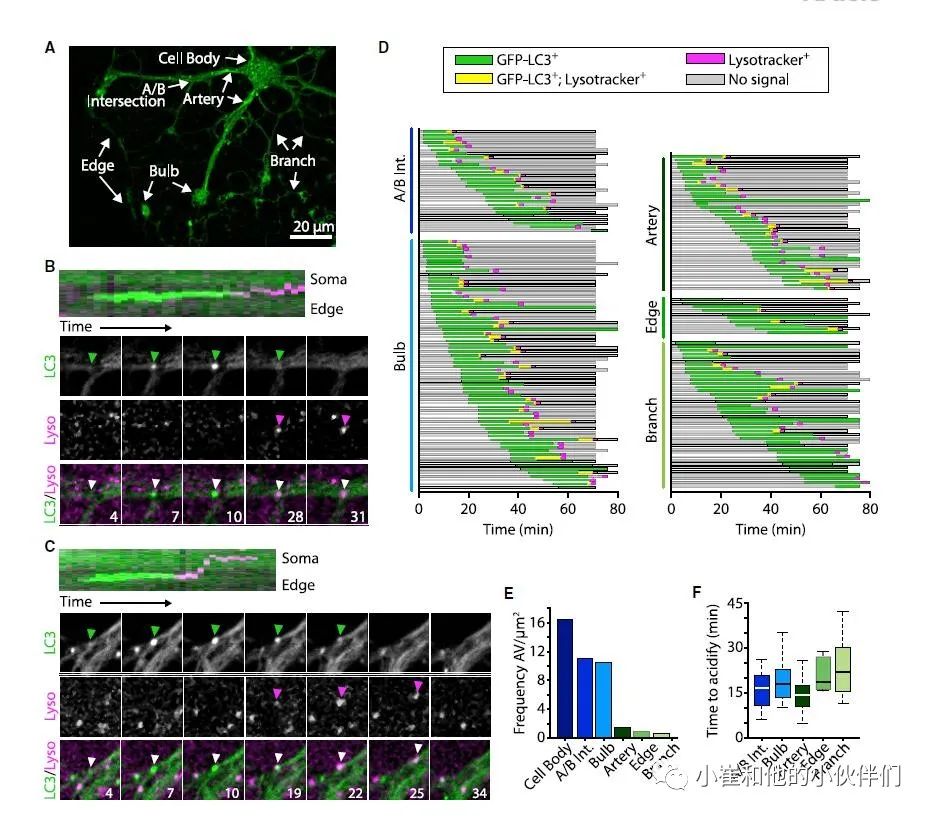 图 3. 自噬体的形成和成熟发生在整个成熟的少突胶质细胞中,
图 3. 自噬体的形成和成熟发生在整个成熟的少突胶质细胞中,
可通过活细胞成像选择性地捕获 MBP
髓鞘碱性蛋白是选择性自噬的一种载体
作者接下来试图识别被巨自噬作用捕获的髓鞘蛋白,髓鞘碱性蛋白(MBP)是髓鞘的重要细胞溶质成分,适度的溶酶体抑制导致MBP积累并与内源性LC3和LAMP2共定位,这与它是一种自噬载体相一致(图4A)。接下来,作者培养了巨自噬缺陷的少突胶质细胞来进一步研究这个问题。作者把目光转向Atg5缺失上,以便可以通过TdTomato的表达监测Cre活性(Atg7与 Rosa26基因位于同一染色体上)。
使用这种方法,作者发现 CNPCre在分离的少突胶质前体细胞中是活跃的。与作者团队对 Atg7 的体内发现一致,而Atg5 缺失抑制自噬(图4B 和4C),并且与之前的报告相比,它对体外少突胶质细胞分化没有明显影响。少突胶质细胞裂解物的免疫印迹显示,Atg5的缺失导致MBP在洗涤剂不溶性池中积累,梯度增加,表明 MBP 聚集(图4D),成年 Atg5cKO 小鼠的胼胝体中MBP 水平也更高,与Atg7cKO 小鼠类似,并延伸到皮质层(图4E 和4F)。在培养中,p62和MBP的共染不仅证实MBP+包涵体的存在,而且其中许多包涵体对p62也是阳性的(图4G和4H)。鉴于MBP 本质上是一种无序的蛋白质,具有高度聚集倾向,并且p62与巨自噬的聚集周转有关, 作者接下来测试验证了MBP和p62共定位的假设,因为 MBP 是被选择性自噬降解的。
选择性自噬是自噬载体以适应蛋白依赖方式被转运降解,以允许选择载体。不同的载体可以被选择性地降解,例如线粒体(线粒体自噬)、入侵病原体(细菌自噬)和聚集蛋白(聚集性自噬)。在聚集性吞噬中,衔接蛋白 WDFY3(也称为自噬相关的FYVE 蛋白[Alfy])与 p62一起通过自噬降解聚集的蛋白质。野生型(WT)小鼠白质MBP的免疫共沉淀实验表明,MBP与p62、Alfy和LC3形式I和II处于一个复合物中 (图4I)。对具有轻度溶酶体抑制的少突胶质细胞的超微结构研究显示,在没有大量胞质溶质的双膜结构中存在电子致密聚集体(图4J),这是细胞质对液泡靶向的标志,是哺乳动物聚集的酵母范式。这些数据强化了MBP以与选择性自噬一致的Alfy依赖的方式降解的观察结果。
 图 4. 少突胶质细胞蛋白 MBP 被选择性自噬降解、
图 4. 少突胶质细胞蛋白 MBP 被选择性自噬降解、
完整的膜髓鞘蛋白需要巨自噬才能降解
与MBP一样,免疫荧光显示,溶酶体的抑制增加了跨膜髓鞘蛋白的积累,例如髓磷脂髓鞘蛋白(PLP)和髓鞘少突胶质细胞糖蛋白(MOG)(图5A和5B),并增加了它们与 LC3+ 结构的共定位(图5C),预示其降解需要巨自噬的参与。为了进一步研究这一假设,作者对成年 WT 小鼠的大脑匀浆进行了密度梯度离心,以富集自噬小泡 (AVs)(图 5D)。
运用免疫印迹显示 MBP、MOG、PLP 和髓鞘相关糖蛋白 (MAG) 分馏为 AV 部分(图5E)和预期的富含髓鞘的光膜部分,进一步表明它们需要巨自噬丰富的自噬小泡分离潜在的ER微粒体和内吞结构,作者接下来对部分 F5进行免疫分离,以获得 LC3 阳性的囊泡(图5D),免疫印迹显示,免疫分离进一步富集了髓磷脂蛋白的部分(图5F)。此外,蛋白酶 K (PK) 保护试验证实 MBP、MOG、MAG 和 PLP 与 p62 一起被内化在自噬体中(图5G)。
综上所述,这些数据强烈表明 LC3+ 自噬结构捕获了完整的髓鞘膜蛋白。PK保护试验还进一步强调了MBP在自噬小泡内处于具有高度有序结构的假设:在没有洗涤剂的情况下,新的 MBP+条带以 PK 剂量依赖性方式出现,表明MBP在自噬小泡内存在洗涤剂和 PK 抗性的聚集体。综上所述,这些数据与巨自噬降解髓鞘的细胞溶质和整体膜成分的观察结果一致。
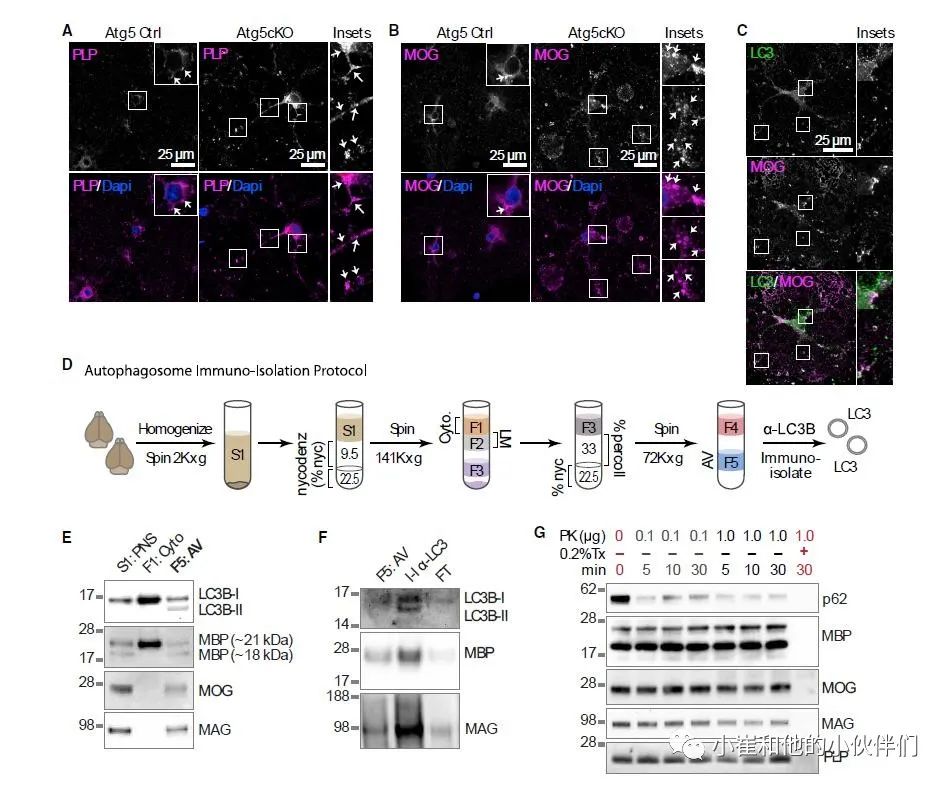
图 5. 完整的膜髓鞘蛋白降解需要巨自噬的参与
自噬内涵体将髓鞘的细胞质和完整膜成分聚集在一起以允许它们被溶酶体降解
接下来,作者通过免疫荧光方法验证了自噬小泡中发现的完整髓鞘膜蛋白的来源(图5)。鉴于这些细胞表面蛋白必须首先通过内吞作用被内化,表明它们通过自噬内涵体的形成被带到 LC3+ 结构中。自噬内涵体通过内吞囊泡与新生的自噬体融合形成,并可能在溶酶体介导的胞体降解之前巩固载体,尤其是在神经元中。在WT少突胶质细胞中,mCLING 阳性结构的一个子集——种标记细胞质膜的荧光标记的可固定探针——与 LC3 共定位,类似于 MOG(图5C),强烈支持在少突胶质细胞中形成自噬内涵体的假说。
同样,免疫荧光显示,mCLING+ MOG+结构在对照 组(Ctrl)培养物中与MBP共定位,表明当自噬作用完好时,内吞和胞质载体可以到达相同的囊泡结构(图6A和6B)。对 Ctrl少突胶质细胞的超微结构研究还揭示了携带髓鞘类载体的自噬内涵体结构的存在:该结构类似于双膜囊泡,与充满髓鞘类膜的单膜囊泡融合(图6C)。还发现了充满电子致密物质的单层结构和被认为是自溶酶体的膜结构(图6D)。
尽管在 Atg5 Ctrl 和条件敲除 (cKO) 培养物中,整个少突胶质细胞中都观察到 mCLING+MOG+结构的内化(图 6A和6B),但在 Atg5cKO 的少突胶质细胞中,MOG 和 PLP 在整个细胞内积累(图5A 和5B)。mCLING+MOG+囊泡很少与内吞载体共定位,例如转铁蛋白受体 (TfR),它们不会在 Atg5cKO 细胞中积累。这与永生化细胞中的数据一致,表明完整膜的髓鞘蛋白不会像TfR那样经历内吞作用。这些数据表明,尽管通过内吞作用内化,但整体的髓鞘膜蛋白仍然需要自噬才能降解。由于髓磷脂膜特别富含胆固醇和鞘脂,并有 PLP 等蛋白质与胆固醇相互作用,自噬内涵体可能以网格蛋白介导内吞的方式到达溶酶体。
为了进一步研究这一点,作者利用GFP-LC3+少突胶质细胞的活细胞成像,通过 GFP-LC3+ 自噬体与mCLING+ 核内体的融合来捕获自噬内涵体的形成(图6E–6G),与mCLING的结合导致 mCLING+结构在整个细胞内广泛分布,与 GFP- LC3+ 囊泡的分布相当,但数量更多。此外,作者观察到 GFP- LC3+ 囊泡在整个细胞中与mCLING+ 结构融合,平均约 13% 的GFP-LC3囊泡显示出这种融合事件,数据摘要如图 6G 所示,这些数据表明,自噬内涵体在少突胶质细胞中形成,从而可以将整体的膜蛋白运输到自噬体中。作者的研究表明了一个自噬内涵体协调自噬和内吞作用的模型,这对于溶酶体介导的这种复杂和高度适应的髓鞘降解所必需的。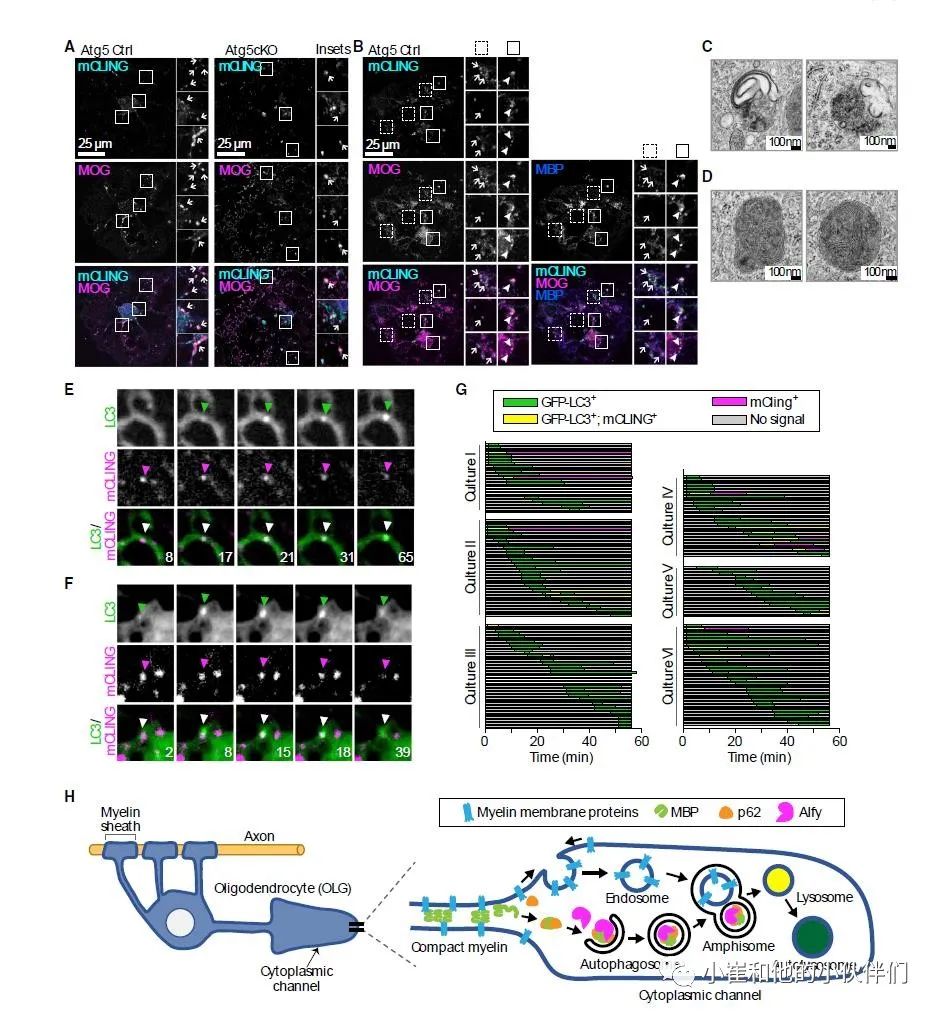
图 6. 髓磷脂相关整合膜蛋白(如 MOG)通过胞吞作用内化,
后形成自噬内涵体进行降解
少突胶质细胞中巨自噬的失活导致行为功能障碍和神经退行性病变
作者团队发现少突胶质细胞巨自噬的失活阻碍了成年大脑中髓鞘的更新,并导致随着动物年龄的增长髓鞘增厚(图 1)。与衰老相关的表型一致,尽管两者都不像Atg7cKO 和 Atg5cKO 小鼠表现出发育缺陷的迹象,但随着它们的成熟,它们从大约 8 个月大开始出现震颤和运动缺陷,然后过早死亡(图7A-7C)。免疫荧光显示,老年自噬缺陷小鼠的皮层出现髓鞘蛋白的聚集、髓鞘变性和神经胶质变性(图7D-7G)。
在超微结构水平上,髓鞘碎片和异常髓鞘形态,包括异常扩大和移位的内环,也很普遍(图7I)。许多这些超微结构异常先前已被证明会破坏轴突传导,这可能解释了为什么会出现运动症状。异常的髓鞘形态通常伴有轴突小球的存在(图7I),这是轴突退化的迹象。在这些大脑中,神经元的死亡也很明显,正如在老年 Atg7cKO(图7G和7H)和 Atg5cKO小鼠的神经元中存在活化的caspase-3,这一点非常值得注意,因为Cre在胼胝体中表达,但在皮质区域没有异位表达。
综上所述,这些数据强烈表明,通过自噬内涵体协调降解来调节髓鞘厚度的能力的丧失会破坏神经回路,导致行为障碍,并最终导致死亡。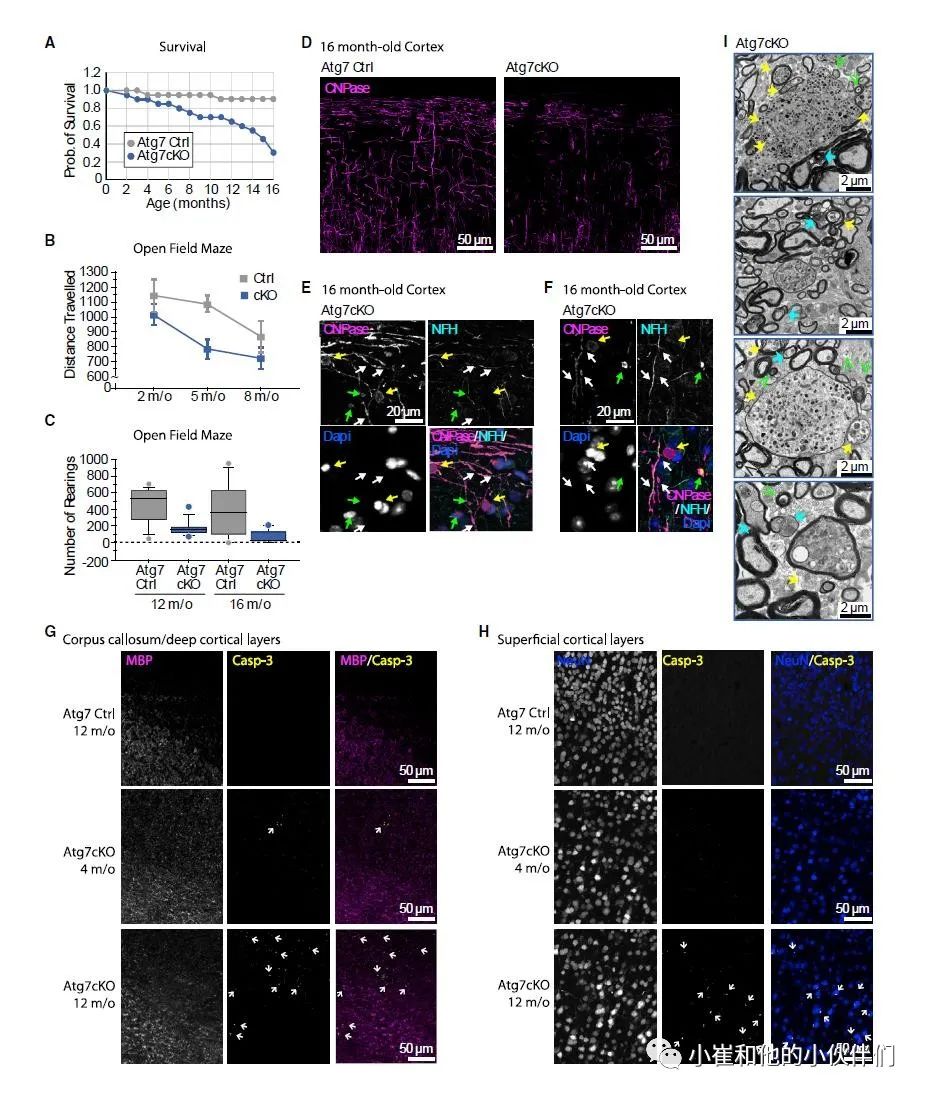 图 7. 少突胶质细胞自噬的失活导致行为障碍、神经退行性病变和死亡
图 7. 少突胶质细胞自噬的失活导致行为障碍、神经退行性病变和死亡
结论
综上所述,当评估巨自噬功能障碍如何导致神经退行性变时,我们必须更好地了解细胞内事件如何维持中枢神经系统中不连续的、多样的和高度适应的细胞类型的稳态。在这里,作者展示了少突胶质细胞如何使用巨自噬这条通路来满足髓磷脂周转的独特需求,以及这条通路的中断如何产生细胞自主和非细胞自主的后果。此外,作者证明了成人大脑的髓鞘可以被细胞自主降解,这使得我们可以对髓鞘的潜在变化如何影响神经回路的形成和维持进行更加深入的思考。
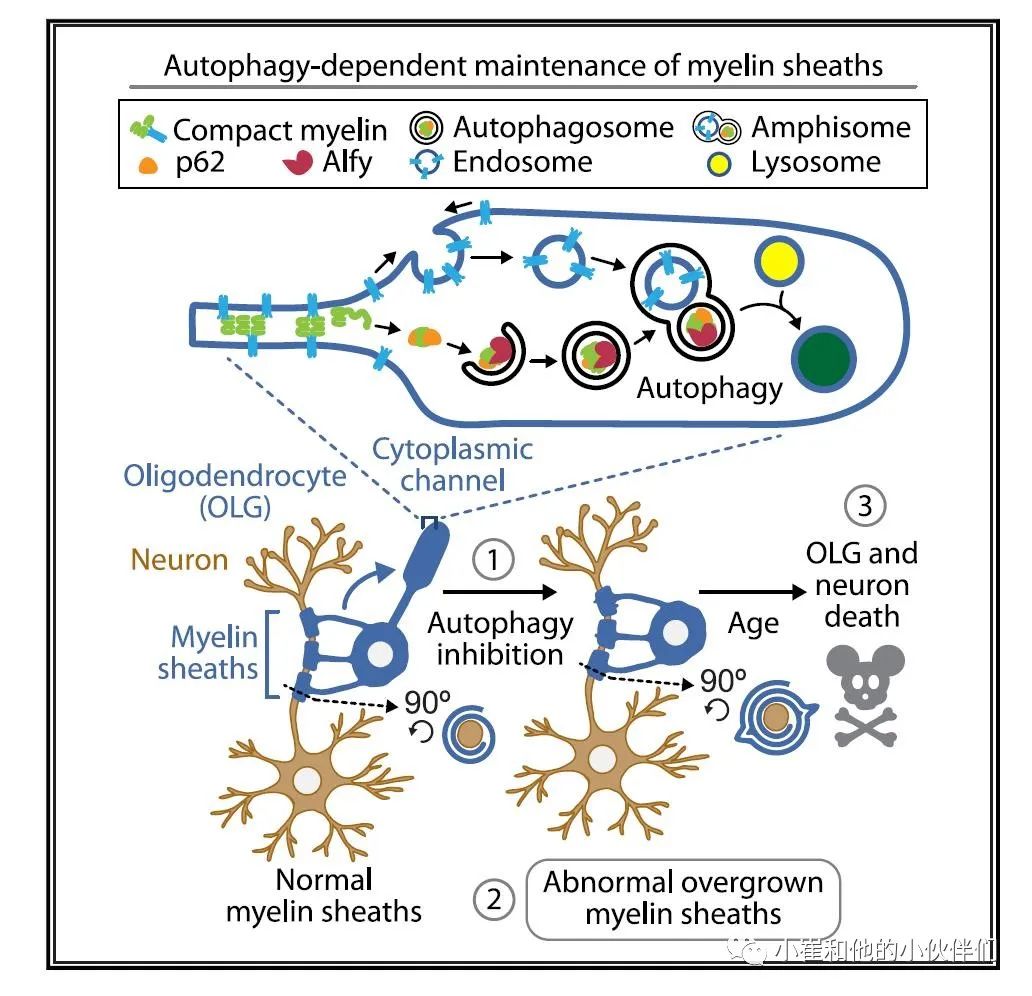 少突胶质细胞组装髓磷脂,并支持其髓鞘化轴突。
少突胶质细胞组装髓磷脂,并支持其髓鞘化轴突。

