五大新兴科学研究方法和项目,你认识几个?|《自然》技术特写
时间:2021-07-12 17:02:06 热度:37.1℃ 作者:网络
回顾迅速崭露头角的杰出研究工具和项目,我们可以发现一些共同的成功途径。
当被问及专长时,Kaihang Wang的回答很干脆:“手艺人”。毕竟他在加州理工学院(California Institute of Technology)的大部分工作都与造东西有关,尽管不是用锤子和钉子。Wang的团队开发了分子工具,包括一个系统——生物学家可以通过编程,将长的合成DNA链转入细菌细胞[1]。再度思考过后,Wang给出了一个更科学的回答:合成生物学或基因组工程。“从根本上说,我们所有努力主要由一个基本目标推动,那就是创造生命”,他说。
和Wang一样,当手头的工具不足时,许多生物学家会跨学科寻找材料、合作者或不同的方法。这促成了全新命名的方法或联盟,如“膨胀显微成像(expansion microscopy)”或“基因组编写计划(Genome Project-write)”。其中一些方法或联盟由于其技术能力及显赫的名声而在科学家中引起轰动。

即将到来:“人类细胞图谱”。来源:改编自Getty。
科罗拉多州立大学研究科学修辞学的Erika Szymanski表示,为一个领域或工具取个琅琅上口的名字,可以为研究者创建出探索的概念框架。“就像显微镜限制了我们用它能看到什么,我们只能‘看见’那些有名字的东西,”她说,“尝试以新框架来思考工作有时会很有成效,因为它开辟出空间,让我们可以想象新的可能性。”
在本文中,《自然》探索了过去15年中5项著名的技术。有些已经开辟了新的研究领域或取得了资金资助;有些加强了全球合作,或者在研究中发现了不同于最初意图的新目标。无论是揭示了细胞功能,催生了公司和疗法,还是在疫情期间为公共卫生决策提供了信息,这5项技术都在科学史上留下浓墨重彩的一笔。
表观转录组学
与基因组DNA一样,信使RNA可以携带改变其功能或命运的化学标记,例如甲基或糖基。这种修饰并不统一,并且有发现表明,某些mRNA高度甲基化而其他mRNA没有,指向了这些标记的生物学作用。2012年,威尔康奈尔医学院(Weill Cornell Medical College)的RNA生物学家Samie Jaffrey等开发了一种方法来识别普遍存在于转录组(细胞或生物体中转录出来的所有RNA)中的特定mRNA甲基化标记,命名为m6A[2]。
该研究的共同作者Christopher Mason也在威尔康奈尔医学院工作,他创造了“表观转录组学”这一术语来解释该团队的假设,即甲基标记调节mRNA转录本的活性,从而表明为什么蛋白质水平并不总是与编码它们的转录本的丰度相匹配。“这可能是遗传编码的新层面,这一点很吸引人。”Jaffrey说。新名称使其他人更容易理解这个概念。
几年下来,表观转录组学已经发展成一个独立的领域,有专门的资金、会议和合作需求。西班牙巴塞罗那基因组调控中心 (Centre for Genomic Regulation,CRG) 的RNA生物学家Eva Maria Novoa Pardo说:“在某种程度上,一个新词的创造引领了整个科研群体的出现。”
Jaffrey和Mason的早期方法是使用m6A抗体来分离长为100-200个核苷酸的修饰RNA片段,然后他们通过测序对其进行鉴定。后来,该团队将抗体与底物交联,然后沉淀抗体结合的RNA片段以精确定位甲基化位点,从而生成第一个单核苷酸水平的甲基化mRNA图谱。这有助于识别另一类携带修饰的分子,称为核仁RNA[3]。“我们现在开始认同一个想法:m6A的一个主要功能是标记RNA以实现快速周转”,Jaffrey说,这对细胞改变和适应环境的能力至关重要。
随后科学家开发了可以在特定序列上切割非甲基化RNA的酶。开发者、以色列魏茨曼科学研究所(Weizmann Institute of Science)RNA生物学家Schraga Schwartz利用该工具,不仅能检测特定位点是否被修改,还可以检测携带甲基化基序的转录本的百分比。当Schwartz等将其应用于整个转录组时,他们发现基于抗体的技术遗漏了近75%的修饰位点,表明其敏感性有限[4]。“这个结果令人惊喜,”他说,“以前就一种,现在有了两种方法,我们看问题更全面了。”
如今,表观转录组学研究人员可以使用纳米孔测序仪直接读取修饰过的 RNA。与传统测序仪需要先通过逆转录将RNA转化为DNA不同,这些仪器将RNA分子通过蛋白质纳米孔并产生特定的电流,然后解码电流信号以获取RNA序列。过去,解码电流信号的测序算法经常误读甲基化的m6A核苷酸。因此,2019年Novoa等人设计了一种算法(今年早些时候有更新[5]),使用这些错误来预测哪些位点携带甲基化核苷酸。“有可能对天然RNA进行测序(而无需先将其逆转录成DNA),为转录组开辟了无偏差的图景”,她说。
人类细胞图谱
2003年人类基因组测序的完成,以及研究单细胞的新工具的出现,让科学家开始畅想是否可以对每个人类细胞的独特位置、行为和发育进行绘图。英国维康桑格研究所(Wellcome Sanger Institute)遗传学家Sarah Teichmann和美国南旧金山基因泰克(Genentech)的计算生物学家Aviv Regev就是其中两位。
2016年底,Teichmann、Regev等聚在一起讨论这个想法。人类细胞图谱计划(Human Cell Atlas)由此诞生,这是一个使用单细胞途径绘制每个人类细胞、组织和器官的结构、遗传学和生物学的项目。该小组强调开放、协作的方法:任何人都可以参与,并且该联盟使用广泛的分子和计算方法收集信息。
“没有什么金标准技术可以实现所有目的,”在CRG 研究单细胞测序技术并领导该联盟标准和技术工作组的Holger Heyn说,“每种方法都有误差。我们整合的技术越多,误差就越少。”
在2020年的一项研究中,Heyn等人在一组普通参考样本中比较了13种单细胞RNA测序技术,并根据其发现细胞特异性标记物的能力进行评价[6]。他们发现,结果差异的一个主要来源是样本中细胞的大小。“我们的目标不是比个高下,而是决定通过每种技术能获取哪些信息”,Heyn说。
人类细胞图谱联盟现在在77个国家拥有近2200名成员,他们总共分析了来自14个主要器官的约3900万个细胞,并发表了近80篇文章,而且这些数字还在不断增加。
此外,这些数据还有助于解开COVID-19的奥秘。2020年初,联盟成员汇集了26个已发表和未发表的数据集,以了解冠状病毒SARS-CoV-2如何入侵肺组织。他们绘制了病毒用于进入组织(包括鼻子、嘴巴和眼睛等)的细胞表面受体图[7]。此后,世界各地的研究人员使用该图谱来了解感染过程。Teichmann表示,它甚至有助于为公共卫生决策提供信息,例如要求人们戴口罩的政策。“这场疫情对人类细胞图谱计划来说确实是变革性的,”她说,“它展现了细胞图谱的价值——即使还是早期的、不完整的图谱。”
膨胀显微成像
尽管许多着迷于显微镜分辨率的研究人员专注于打造更好的硬件,但神经科学家Ed Boyden采取了不同的策略。他与麻省理工学院的同事一起,设计了一种称为膨胀显微成像(expansion microscopy)的技术,它可以像给气球打气一样扩大细胞和组织。
该方法将一种称为丙烯酸酯的单体注入样品中。加水会导致单体聚合和膨胀,随着其扩大,细胞组分被推开。早期尝试时细胞会破裂或膨胀不均匀。但通过在聚合前添加酶来软化组织,研究人员可以将小鼠脑组织扩大到原始大小的4.5倍[8]。两年后,该团队将该方法延伸至十几种组织类型,其中一些可以扩大16倍[9]。“能确保物理放大倍数的比例正确,这个技术才有价值,”Boyden说。
今年,Boyden团队利用这个概念来定位组织中的特定RNA,这是一个称为空间转录组学的子领域。他们首先扩展了小鼠脑组织的一部分,然后对锚定的RNA进行了原位测序[10]。
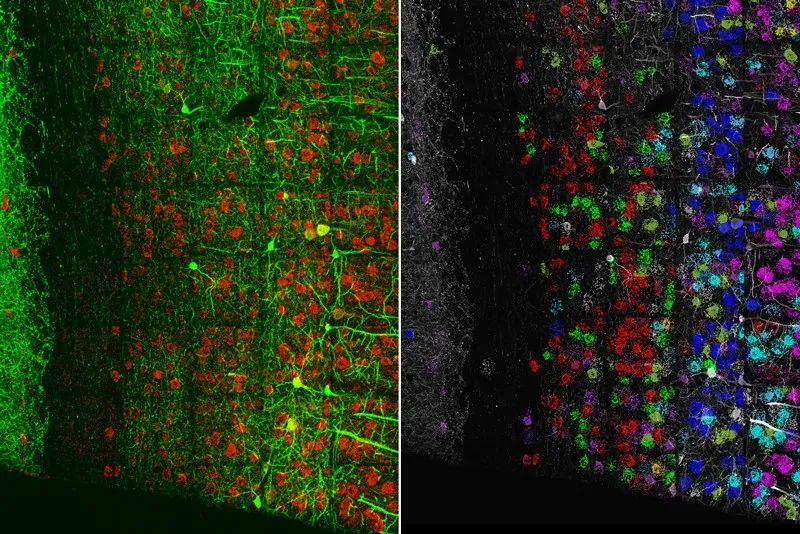
膨胀显微成像联合RNA测序(左)共同揭示了小鼠视觉皮层神经元的结构(右)。 来源:S. Alon et al./Science
德国马克斯普朗克脑研究所(Max Planck Institute for Brain Research)的神经科学家Erin Schuman研究蛋白质在名为突触的神经细胞连接处如何合成,长期以来他一直依靠银染色等间接方法来可视化此过程。Schuman想直接在突触中看到新合成的蛋白质。但突触是由长而细的纤维形成的,这些被称为轴突的纤维缺乏良好的分子标记。“它们其实是那种最难研究的东西”,她说。
通过膨胀显微成像技术,Schuman团队第一次看到,几乎所有的轴突末端都有合成新蛋白质的机制[11]。“它确实帮我们以高置信度接触突触,并进行高通量分析”,她说。
斯坦福大学(Stanford University)生物工程师Bo Wang使用该工具创建了一张高分辨率图像,展示了常见肠道病原体沙门氏菌如何与人体细胞相互作用。在优化“软化”步骤时,Wang和同事发现该方法可用于测量细菌细胞壁的硬度。这个坚硬的外层,是该病原体对抗生素和宿主防御的关键。测量微型物体的机械特性很困难,但膨胀显微成像技术帮助团队测量了单个批次中数千个细胞壁的强度,以了解细菌如何对宿主防御机制做出反应[12]。“类似的策略可以帮助回答植物、真菌和许多不同物种的生理问题”,Wang说。
神经彩虹
2007年,由哈佛大学神经科学家Jeff Lichtman和Joshua Sanes领导的团队开发出一种方法来区分小鼠大脑中纠缠的神经元[13]。研究人员构建了一个系统,其中编码少数荧光蛋白的基因由神经元特有的调节序列控制,该序列两侧是标签,标签将引导重组酶对这些荧光基因进行随即表达。细胞会得到基因“盒”的多个副本,当研究人员激活识别重组标签的蛋白质时,它会将这些基因改组为各种随机组合,并表现为如彩虹般的荧光。他们称此工具为脑虹(Brainbow)。
Gabriel Victora回想起自己在纽约大学(New York University)攻读研究生时,对那些如万花筒般绚烂的大脑图片大感震撼,每个细胞颜色都不一样。但Victora的研究集中于生发中心(淋巴结的一种微观结构,免疫细胞在此分裂和生长)。“我们没有立即想到可以用这项技术,”如今已是纽约市洛克菲勒大学(Rockefeller University)免疫学家的Victora说,“我记得当时在想,‘可惜是那是在大脑里’。”
Lichtman曾希望标记单个细胞的能力将有助于解决精细尺度的细节问题,例如大脑中的突触连接。但是小的细胞结构荧光分子少,产生的荧光信号亮度不够——通常都太暗了没法用。Lichtman表示,他对结果感到失望,此后转向了诸如连续切片扫描电子显微镜之类的技术,在这种技术中,一块组织被重复成像、切削、再次成像,以绘制神经连接图。“你得为这项工作找到合适的工具,在这种情况下,Brainbow不够用,”他说。
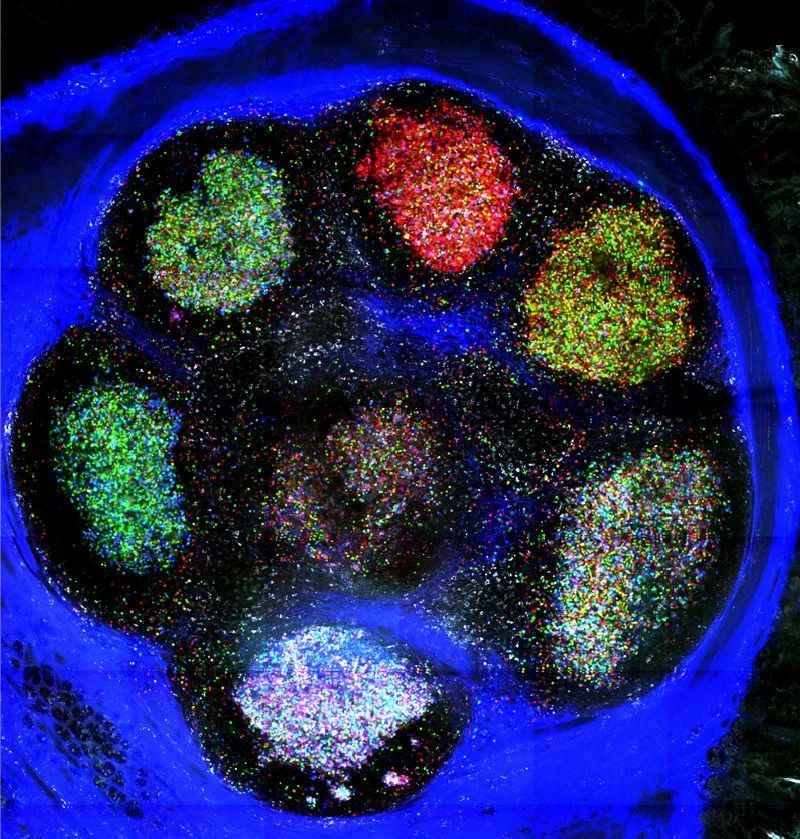
脑虹标记的生发中心。 来源:Carla Nowosad
Lichtman确实使用Brainbow在周围神经系统做了实验,其中细胞相距较远,因此微弱的荧光也可以观察到。其他团队已经针对不同生物调整了工具——例如果蝇大脑的 Flybow和斑马鱼组织的Zebrabow。Brainbow与膨胀显微成像技术相结合,使研究人员能够检查哺乳动物组织中的细胞形状和连通性[14]。
而在Victora那里,有一种名为Confetti的小鼠模型将脑虹技术扩展到了非神经元细胞,这重新点燃了他对Brainbow的兴趣。在淋巴结的生发中心内,成群的B细胞分泌不同抗体,并彼此竞争。大多数生发中心保持着抗体分子的多样性。但Victora团队发现,在5-10%生发中心内,能产生高亲和力抗体的B细胞数量可以迅速超过其它B细胞,并接管生发中心[15]。通过Brainbow追踪这些“克隆爆发(clonal burst)”的研究人员在第一次标记细胞时,看到生发中心的所有细胞都呈现不同的颜色。然后,当一个优势克隆接管时,它的后代——所有这些都与亲代细胞具有相同的颜色——将生发中心从彩色变为单色。他说:“Brainbow非常清楚地显示了B细胞之间这种的分工。”
基因组编写计划
如果科学家能够合成完整的染色体,他们就可以赋予细胞新的功能,更换致病的遗传途径或设计新的实验系统进行研究。但是,染色体合成不能一蹴而就。
2010年,研究人员拼凑出第一个细菌的合成基因组[16]。他们将细菌DNA改造成短片段,再将它们拼接在一起,然后一次一个片段地交换一部分染色体,直到原始DNA完全被合成对应物所取代。加州理工学院的Wang说,自从第一次尝试以来,这个过程基本保持不变。尽管在细菌和酵母方面取得了显著进展,但该技术从未拓展至基因组更复杂的生物。因此,在2016年,研究人员宣布了基因组编写计划(Genome Project-write),旨在合成复杂的基因组,包括人类的基因组。
该项目(Nature 557, 16-17; 2018)启动时雄心勃勃,由于资金和技术的双重挑战,后面却不得不降低期望,专注设计一种能抵抗病毒的人类细胞系。但这种规模的DNA合成仍然很难,设计编码新功能的遗传线路也一样。麻省理工学院的合成生物学家Christopher Voigt表示,目前,这类工作很大程度上仍属于个别研究员或小团队的单打独斗。如果想要大规模基因组合成变得可行,那么这个过程必须改变。“这就像单人造飞机,从设计到组装什么都做,”他说,“这说明了我们距离在基因组这个规模上做设计有多遥远。”
尽管如此,Wang认为这个崇高的目标仍然可以推动领域向前发展。“合成全基因组的动机推动了技术的发展。这是一个良性循环:一旦我们有了工具,它就会使基因组合成更加可行,人们也会将更多资源投入该领域。”
参考文献:
1. Fredens, J. et al. Nature 569, 514–518 (2019).
2. Meyer, K. D. et al. Cell 149, 1635–1646 (2012).
3. Linder, B. et al. Nature Methods 12, 767–772 (2015).
4. Garcia-Campos, M. A. et al. Cell 178, 731–747 (2019).
5. Begik, O. et al. Preprint at bioRxiv https://doi.org/10.1101/2020.07.06.189969 (2021).
6. Mereu, E. et al. Nature Biotechnol. 38, 747–755 (2020).
7. Sungnak, W. et al. Nature Med. 26, 681–687 (2020).
8. Chen, C., Tillberg, P. W. & Boyden, E. S. Science 347, 543–548 (2015).
9. Chang, J.-B. et al. Nature Methods 14, 593–599 (2017).
10. Alon, S. et al. Science 371, eaax2656 (2021).
11. Hafner, A.-S., Donlin-Asp, P. G., Leitch, B., Herzog, E. & Schuman, E. M. Science 364, eaau3644 (2019).
12. Lim, Y. et al. PLoS Biol. 17, e3000268 (2019).
13. Livet, J. et al. Nature 450, 56–62 (2007).
14. Shen, F. Y. et al. Nature Commun. 11, 4632 (2020).
15. Nowosad, C. R. et al. Nature 588, 321–326 (2020).
16. Gibson, D. G. et al. Science 329, 52–56 (2010).
原文以Five trendy technologies: where are they now?标题发表在2021年6月21日的《自然》的技术特写版块上
© nature
doi: 10.1038/d41586-021-01684-7

