Science:小胶质细胞调节阿尔茨海默病和帕金森病的神经退行性变
时间:2022-10-23 12:00:27 热度:37.1℃ 作者:网络

目前观点认为,淀粉样蛋白并不一定是神经退行性疾病中神经元功能障碍和认知衰退的始作俑者。对百岁老人的研究发现,尽管他们脑中富含淀粉样蛋白,但仍有良好的认知表现。流行病学研究提出了认知储备的概念,具有良好认知储备的个体可以更好的适应大脑的病理改变,从而延缓或阻止发病。因此,神经变性疾病研究的挑战之一即是了解这些个体在脑中淀粉样蛋白沉积的存在下,大脑是如何成功地维持正常的神经元功能的。基因研究表明小胶质细胞(大脑内主要的巨噬细胞)可能是调控和维持神经元功能的关键。那么,在AD和PD中,是否存在小胶质细胞-神经元相互作用的受损呢?哪些方面的损害导致了神经退行性变?
为解答上述问题,来自伦敦大学学院的Soyon Hong教授团队2020年10月在Science“Neurodegeneration”版块发表综述,阐述了小胶质细胞引起AD和PD神经元功能障碍的可能途径,题目为“Microglia modulate neurodegeneration in Alzheimer's and Parkinson's diseases”。
1小胶质细胞对神经元功能和健康至关重要
基因组和蛋白质组学证据表明,小胶质细胞类似于其他组织内的巨噬细胞,根据所处生物状态的不同(即大脑区域、年龄、健康和代谢需求)而具有不同的功能。除了免疫功能之外,小胶质细胞对大脑发育至关重要,例如,在大脑发育过程中促进神经元突触的形成。在成人大脑中,小胶质细胞执行多种功能,包括监测神经元活动的变化,调节学习和记忆,以及充当大脑局部的吞噬细胞和损伤传感器。
许多小胶质细胞-神经元的相互作用是由细胞-细胞信号通路介导的,包括嘌呤信号传导、细胞因子、神经递质和神经肽。这些功能通常需要高能量消耗及线粒体正常代谢维持,因此小胶质细胞在急性低血糖状态下表现出代谢灵活性。在AD和PD中,慢性线粒体功能障碍是否会损害小胶质细胞的代谢适应能力从而能够监测和调控神经元功能?在神经元线粒体损害和神经退行性病变中,胶质细胞会积聚脂滴,调节巨噬细胞的功能。并且,小胶质细胞中的脂滴积累和年龄增长相关,那么,老化的小胶质细胞中脂代谢的异常是否是AD和PD的易感性基础呢?目前已发现的多种AD和PD危险因素,包括TREM2、载脂蛋白E (ApoE)、GBA1和硬脂酰辅酶a去饱和酶(SCD),被发现可以调节脂质代谢、溶酶体途径和小胶质细胞代谢适应性,似乎进一步支持了这一假设。
2小胶质细胞介导AD突触丢失
在AD中,突触丢失和功能障碍具有区域特异性及早期发生的特点,并与认知障碍密切相关。β-淀粉样蛋白(Aβ) 寡聚体和/或tau被发现在突触中聚积和引起突触功能障碍和丢失。目前半数以上的AD遗传危险因素是由髓样细胞表达的。
多项AD动物模型研究表明,突触存在多种神经免疫信号通路的失调,包括经典补体级联通路、TREM2、磷脂酰丝氨酸(PtdSer)和载脂蛋白E(ApoE) (图1)。那么,突触上局部病理蛋白的积累是否会扰乱对突触健康至关重要的神经元-胶质细胞之间的相互作用呢?例如,积聚在突触上的病理性Aβ或tau可以上调小胶质细胞中的C1q,并促进突触中的补体激活和随后的小胶质细胞吞噬。用基因或基于抗体的方法阻断AD小鼠模型中经典补体级联通路的激活已被证明可以保护突触,避免突触丢失、功能障碍和记忆丧失,这提示小胶质细胞-突触通路可能是一个潜在的治疗靶点。在一开始,这种通路可能是一个有益的过程,即小胶质细胞吞噬异常的突触,但是慢慢的,小胶质细胞出现功能失调,并损害它曾试图挽救的神经元。这种小胶质细胞吞噬异常突触的机制被称为是小胶质细胞-突触修剪机制。由于包括突触修剪在内的许多小胶质细胞功能似乎是活动依赖的,因此早期AD小鼠模型中观察到的神经元过度活跃可能提示小胶质细胞异常吞噬突触的存在。深入了解调控修剪的途径和引导小胶质细胞吞噬突触的特定信号,有助于确定认知功能下降的潜在治疗靶点以及发现候选生物标记物来量化与突触丢失相关的小胶质细胞功能障碍。
图1. AD小胶质细胞中补体介导的突触丢失
另一生物学和治疗学上重要的问题是:小胶质细胞是否会靶定和清除特定的突触?人类和小鼠AD脑中突触小体的蛋白质组学研究发现了突触的线粒体功能障碍。然而,补体因子(包括C1q和C3b)是否针对特定的突触(即功能障碍和/或受损)尚不清楚。神经元-胶质细胞相互作用中的脂质信号可能是一个关键的决定因素。例如,在小鼠海马中,TREM2(一种小胶质细胞的关键损伤传感器)已被证明参与突触发育过程。TREM2的一种配体是暴露在神经元膜外的Ptdser。因此,小胶质细胞TREM2可能通过PtdSer信号感知AD中受损的突触。在脑中含量丰富的神经节苷脂也被认为与小鼠大脑中Aβ诱导的突触功能障碍相关。结合Aβ的GM1神经节苷脂在AD早期大脑的神经元膜上富集。此外,抗GM1神经节苷脂抗体可以将补体固定在神经元膜上,并且在AD模型中使用靶向C1q抗体能改善小鼠体内抗神经节苷脂抗体介导的神经元损伤。这些研究结果提出了一个新的问题,即神经节苷脂是否与AD的突触丢失和补体介导的小胶质细胞对突触的吞噬有关。
此外,在神经免疫和脂质化途径通路中,需要重点关注的是ApoE。既往研究表明,星形胶质细胞ApoE和小胶质细胞突触修剪之间可能存在联系:星形胶质细胞吞噬突触的速率受ApoE等位基因调节,并且与正常突触可塑性相关。这一速率在衰老过程中逐渐降低,使得突触对补体介导的小胶质细胞吞噬产生易感性。值得注意的是,ApoEe4可以增加人类AD大脑突触中的病理性Aβ。最近研究发现,ApoE能结合C1q并调节经典补体级联的激活。以上结果表明,ApoE在星形胶质细胞-神经元-小胶质细胞突触的相互作用中具有重要作用,尤其是在AD中ApoE的特异性失调与胆固醇等脂质代谢途径紧密相关。
TREM2和ApoE是迟发性AD的两个主要危险因素,对这两个因素的研究表明:小胶质细胞的脂质代谢异常是反映大脑免疫系统对淀粉样蛋白慢性堆积敏感性的表现。例如,TREM2缺陷的小胶质细胞会出现慢性脱髓鞘并不能正常代谢脂质。此外,TREM2似乎是主要脂质转运体ApoE的关键调节因子。研究发现,ApoE可将过度活跃的神经元中的过量脂质转运到星形胶质细胞的脂滴中,并在那里进行代谢,这表明ApoE可以改善神经元过度活跃引起的脂质毒性。目前需要明确的是,星形胶质细胞和小胶质细胞中淀粉样蛋白相关的神经元过度活跃和脂质代谢之间的关系如何,以及这种关系为何在老年大脑或具有AD风险基因突变(如TREM2)的大脑中减弱。
3PD胶质细胞:αSyn毒性的调节剂
PD病理改变通常伴随着星形胶质细胞和小胶质细胞中神经元蛋白α突触核蛋白(αSyn)的明显积累,这也是最近被认为是帕金森病小鼠模型的突出特征。此外,改善小胶质细胞-星形胶质细胞的交互作用可以减轻αSyn聚集模型中的PD样病理。这些研究表明胶质细胞在吸收和介导αSyn的神经毒性中起直接作用。值得注意的是,神经吞噬作用——神经元的小胶质细胞吞噬作用——在PD中通过小胶质细胞内神经黑色素的积累得到了证明。PD组织中的突触富含病理性αSyn聚集体,这可能提示PD中存在类似于AD中观察到的突触吞噬机制。然而,补体和小胶质细胞是否介导PD的突触丢失目前尚不清楚。
从遗传学的角度看,PD和小胶质细胞之间的联系并不像AD和小胶质细胞之间的联系那么明显。家族性突触核蛋白病可能与总神经元αSyn的表达水平有关,然而,在散发性帕金森病中,神经退行性变与某些生物活性形式的αSyn密切相关,而与αSyn的总水平无关。此外,三种突触核蛋白病——PD、路易体痴呆(DLB)和多系统萎缩(MSA)——都以淀粉样αSyn负荷为特征,但它们明显表现出不同的脑区特异性蛋白积聚和神经元功能障碍模式。这种显著的区域特异性αSyn扩散模式被认为是由特定细胞外αSyn聚集体或病株的朊病毒样扩散诱导的,类似于朊病毒疾病。PD的遗传学研究表明,鞘脂代谢中有许多疾病遗传危险因素。PD的危险基因GBA1、SMPD1、GALC、ASAH1、CTSD、SPTLC1和SLC17A5参与了溶酶体的降解过程,提示对αSyn聚集体的降解功能失调可能是疾病发生发展的重要机制。
那么,胶质细胞是否参与阻止或促进细胞外αSyn聚集体在不同大脑区域的传递,从而导致突触核蛋白病的区域特异性呢?答案是肯定的。细胞外αSyn聚集体唯一已知的摄取受体LAG3主要由小胶质细胞表达。此外,在最近的突触核蛋白病模型中,破坏小胶质细胞对细胞外αSyn的清除会导致多巴胺能神经元功能失调。这些研究表明,胶质细胞的摄取和加工是调节αSyn活性的关键。因此,虽然胶质细胞在生理环境中可以作为神经元清除蛋白错误折叠的场所,但在疾病中胶质细胞可能是一把双刃剑:胶质细胞对无毒αSyn的摄取和加工,实际上可能是通过自噬以及有缺陷的溶酶体降解产生特异性毒性αSyn的过程(图2)。鞘脂代谢失衡介导的小胶质细胞对细胞外αSyn的病理修饰可能是导致PD、DLB或MSA慢性αSyn功能障碍的关键决定因素。
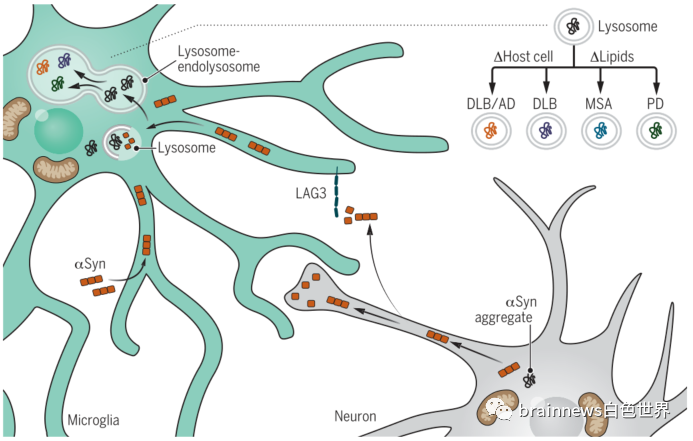
图2.神经胶质细胞在PD中作为αSyn毒性的调节器
4帕金森病的脑外观察:肠道中的巨噬细胞-神经元信号
最新的临床前和遗传学数据表明,在PD病理中,肠道神经系统(ENS)-所谓的肠脑-可能与PD病理有关。研究者已经在ENS中观察到了αSyn聚集,并认为αSyn是以细胞到细胞跨突触的方式从肠道扩散到大脑。切断小鼠迷走神经干发现可以防止病理性的αSyn传入大脑,这表明迷走神经是一条潜在的传递αSyn的管道。值得注意的是,肠道注入的Syn不仅诱导了在肠道神经元的αSyn磷酸化,也刺激了CX3CL1和CSF1的产生,这是与肠道巨噬细胞上的CX3CR1和CSF1R结合的配体。最近的一项研究发现了ENS中一种特定类型的组织驻留巨噬细胞,它类似于大脑中的小胶质细胞,寿命很长,对神经元存活和肠道功能很重要(图3)。这些驻留在ENS上的肠道巨噬细胞表达高水平的转录体,包括Gba1和Lrrk2,这些转录体涉及囊泡运输和内溶酶体途径。LRRK2基因突变是常染色体显性PD的常见原因;然而,LRRK2究竟如何导致Syn病理和PD样症状目前尚不清楚。值得注意的是,缺乏Lrkk2的巨噬细胞显示出更高的蛋白水解活性并表达更高水平的溶菌酶,这表明LRRK2调节溶酶体功能和吞噬体成熟。此外,LRRK2与肌动蛋白重塑因子WAVE2相互作用,调节巨噬细胞的吞噬功能。因此,评估肠道巨噬细胞如何在PD中受到影响并沿肠脑轴调节病理性αSyn的清除,对PD的诊断和治疗意义重大。
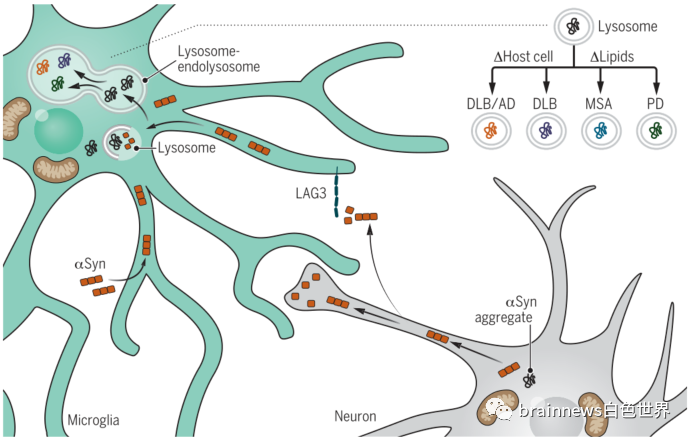
图3.帕金森病患者沿肠-脑轴的巨噬细胞-神经元交互作用
5展望
在AD和PD中,大脑和肠道中的巨噬细胞不能感知功能失调的神经元,这可能会导致神经元稳态和正常功能的病理性破坏。来自患者组织和动物模型的研究明确指出,胶质细胞不仅仅是淀粉样蛋白的吞噬细胞,它们还管理和调节神经元的健康。因此,发展监测活体脑中胶质细胞-神经元交互作用的方法,并评估与疾病相关的通路,对于神经退行性疾病的早期诊断和治疗尤为重要。
神经炎症是有益或有害,需要具体问题具体分析。例如,经典的补体级联反应有助于减少淀粉样蛋白,但也会介导突触丢失。增强TREM2活性可能对存在淀粉样蛋白聚积的脑组织有益,但对存在神经纤维缠结的脑组织则没有明显作用。因此,对于慢性的多因素的疾病,如AD和PD,我们在评估功能和影响的同时也需要仔细考虑具体的机体状态。在疾病的不同阶段使用不同的策略,可以靶向特定的生物过程达到有效的治疗目的。
